Towards quantification of vocal imitation in the zebra finch
|
|
|
- Verity Horn
- 6 years ago
- Views:
Transcription
1 J Comp Physiol A (2002) 188: DOI /s ANALYSIS OF SONG DEVELOPMENT O. Tchernichovski Æ P.P. Mitra Towards quantification of vocal imitation in the zebra finch Received: 18 February 2002 / Revised: 16 August 2002 / Accepted: 5 September 2002 / Published online: 20 November 2002 Ó Springer-Verlag 2002 Abstract The transition from an amorphous subsong into mature song requires a series of vocal changes. By tracing song elements during development, we have shown that the imitation trajectory to the target could not be predicted based on monotonic progression of vocal changes, indicating an internal component that imposes constraints on song development. Here we further examine the nature of constraints on song imitation in the zebra finch. We first present techniques for identifying and tracing distinctive vocal changes, and then we examine how sequences of vocal change are expressed and coordinated. Examples suggest two types of constraints on song imitation, based on the nature of the temporal context. Developmentally diachronic constraints are imposed by sequential dependencies between vocal changes as a function of developmental time, whereas developmentally synchronic constraints are given by the acoustic context of notes within the song. Finally, we show that the tendency of birds to copy certain sounds in the song model before others might be related to such constraints. We suggest that documenting the full range of distinctive vocal changes and the coordination of their expression would be useful for testing mechanisms of vocal imitation. Keywords Birdsong Æ Development Æ Sound analysis Æ Vocal learning Æ Zebra finch Abbreviations BOS bird s own song Æ DVC distinctive vocal change O. Tchernichovski (&) Department of Biology, The City College of the City University of New York, 138 and Convent Ave, New York NY 10031, USA ofer@ccny.cuny.edu Tel.: Fax: P.P. Mitra Department of Theoretical Physics, Bell Laboratories, Lucent Technologies Introduction At about 30 days post hatch, the juvenile zebra finch starts producing the soft, squeaky sounds of its subsong. Within several weeks, these ill-formed sounds transform markedly, as the bird masters the complex sounds of an adult male s tutor song. This process exhibits several stages (Immelmann 1969) and requires auditory feedback (Konishi 1965), suggesting that the bird compares the sounds of its own song (bird s own song, BOS) to the auditory memory of a song model it is attempting to imitate (Brainard and Doupe 2000). Some aspects of this sensory-motor process can be studied on a behavioral level: observations of song development show that the bird can approximate a song model via a generative procedure, by transforming the acoustic features of sounds (Thorpe 1958; Immelmann 1969; Clark et al. 1987; Tchernichovski et al. 2001). In addition, the bird can approximate model sounds by narrowing its vocal repertoire via a selective procedure so as to eliminate sounds that do not match a template (Marler and Nelson 1992). Vocal changes, however, are not fully determined by the acoustic error between the BOS and the model, since there is also an internal component of song development that must be taken into account: First, some vocal changes are hardwired and occur regardless of training (e.g., in socially isolated birds; Price 1979), and second, even vocal changes that are specific to the imitation of a particular model are sometimes counterintuitive, such that the imitation trajectory appears to be indirect (Tchernichovski et al. 2001). In this study, we take the behavioral analysis one step further, and make a preliminary attempt to atomize the generative processes underlying song learning in a few simple cases. We then explore the elementary units of vocal changes and examine sequences of vocal changes to uncover constraints that shape (and could potentially predict) imitation trajectories. To gain statistical power for detecting interactions between environmental and

2 868 internal factors during song imitation, we limit our analysis to the imitation of a single song model across different training regimes. Using this approach we can compare vocal changes across birds that copy the same sounds. Materials and methods In this study we combined new data with results obtained and techniques used in several previous studies (Tchernichovski et al. 1999, 2000, 2001; Tchernichovski and Mitra 2002). Most of the methods are already published and are presented here briefly with appropriate references. Animals The data presented here are from young male zebra finches that were raised at the Rockefeller University Field Research Center and were kept under constant (12:12) photoperiod and food ad libitum throughout the experiments. Experimental groups In Figs. 2, 3, 4, and 5) we present data from eight birds that received the same training regime as in an earlier study (Tchernichovski et al. 2001). Young males were raised by their mothers (no adult male present) until 30 days old. Each juvenile was then placed alone in a soundproof chamber that contained a plastic model of an adult zebra finch male (Fig. 1A). Training started on day 43, when we inserted two keys into the training box (Fig. 1B; Adret 1993). Within 1 36 h birds began to peck at either one of the two keys. Key pecking induced the playback of the same short (1.4 s) model song from a tiny speaker placed inside the plastic bird (Fig. 1C). Each playback consisted of two identical repetitions of a single song motif recorded from an adult bird. Each day consisted of two training sessions. During each session we reinforced the first ten key pecks with a song playback. Additional key pecks were allowed, but were not reinforced, so that the overall daily quota of model song that a bird could trigger was at most 40 song motifs (28 s). Training continued until birds were 90 days old, when song development was complete. In Fig. 6(see Results: Statistical analysis of imitation priorities) we present data from three different training groups. Brief training In nine birds the training regime was similar to that described above, except that birds were trained only for 2 consecutive days: keys were inserted to the training box on day 43 post-hatch. Birds were trained for 48 h, starting from the time of the first key peck (which occurred within less than 36 h from the time of inserting the keys). This gave an overall exposure of less than 1 min to song playbacks (four training sessions of 20 song motifs). The keys were then removed and birds were kept socially isolated in the same training boxes until 90 days old, but received no further training. Moderate training Six birds were trained continuously from day 30 and until day 100 post-hatch. The training regime was similar to that described above (20 song motifs per session). Compared to other training regimes tested so far, moderate training gave the most accurate imitation. Data from Tchernichovski et al. (1999). Overtraining Eight birds were trained continuously from day 30 until day 100 post-hatch, reinforcing an unlimited quota of key-pecks, i.e., every key-peck induced a song playback. For reasons unknown, overtraining inhibits song imitation in zebra finches. Data from Tchernichovski et al. 1999). Song recording All songs were recorded digitally with a sampling rate of 44.1 khz and 16-bit accuracy. In the birds represented in Figs. 2, 3, 4, and 5) we recorded a few minutes of song from each bird at least once a day during days after hatching and at least once a week Fig. 1 A The training apparatus included a plastic model, similar in size and coloration to an adult zebra finch, placed in the middle of the box. B Training starts when two keys are inserted to the box. By pecking at either of the two keys, the young bird could induce a song playback from a tiny speaker placed inside the plastic model. C Spectral derivatives (see Materials and methods) of the song model. In subsequent figures specific parts of this model are circumscribed by a red rectangle
3 869 thereafter until day 90. In the three groups of birds represented in Fig. 6 only the mature song was recorded on day 90 post hatch. Data analysis Spectral derivatives Spectral derivatives provide a representation of song that is similar but superior to the traditional sound spectrogram (Fig. 1C). Instead of power spectrum versus time, we perform multi-taper spectrum analysis and estimate spectral derivatives (changes of power) in both time and frequency axes. From time and frequency derivatives we compute directional derivatives which are locally optimized so as to detect frequency contours of any arbitrary angle in time-frequency plan. Directional spectral derivatives are plotted on a gray scale, where negative values are dark and positive values are bright (Tchernichovski et al. 2000). Similarity measurements We measured similarity between songs using the default parameter setting of Sound Analysis 2 software (Tchernichovski et al. 2000; Tchernichovski and Mitra 2002). The procedure can compare any two song motifs (e.g., song model versus BOS) and detect sections of similarity across the two songs based on four song features: pitch, FM, Wiener entropy, and spectral continuity. The similarity indices used here are all percentage of significant similarity, which is an estimate of the proportion of sounds in the song model to which a similar version may be found in the BOS. In the data presented in Fig. 6B, F we present a breakdown of similarity across single notes: to do so, we categorized a note as similar to the model if at least 50% of it was included in a similarity section. Automatic tracing of imitation trajectories To automatically trace imitation trajectories we first compared the mature version of the BOS to the song model. Song sections that were similar to the model were then traced backwards by comparing the mature version to an earlier version (e.g., a day earlier) and so on recursively as long as the procedure could keep track of the similarity. This procedure does not require partitioning of the developing song prior to analysis (the similarity measurement is performed on the entire song, and detects sections of similarity, which are then compared to the entire version of the song in a previous day, and so on). Note that the procedure can only trace relatively short sections of similarity within a song tracing the entire song motif in a single section is rarely possible; in general, the larger the section traced, and the younger the bird, the more likely the procedure to fail (Tchernichovski and Mitra 2002). To quantify the vocal changes from one hour to the next (Fig. 2D) we examined songs at the beginning of each hour and sampled the first 12 renditions of the appropriate syllable. Results Distinctive vocal changes When examining different developmental stages of a song, it is sometimes possible to automatically detect sections of similarity across versions and to align them during an extended developmental time, as illustrated in Fig. 2A. The red rectangle presents a case where the boundaries of a vocal change can be clearly detected. When the scope of a vocal change is limited in both song time and developmental time, we call it a distinctive vocal change (DVC). Figure 2B presents a very simple vocal change that (perhaps not coincidentally) occurred in a bird that produced an inaccurate imitation of the model song. As shown, a single prototype sound of long duration gave rise to two harmonic stacks, similar to those of the song model (sounds c and d in the Fig. 2B). Examining this imitation trajectory from the bottom to the top of the figure shows that on day 52 post-hatch, a localized attenuation appeared at the middle of the prototype. The attenuation then expanded until it became a stop (day 60), splitting the sound into two. This example indicates that syllable boundaries can change during song development, and other examples will show (Fig. 5) that the acoustic type of sounds can also change during song development. It is therefore very difficult to partition the emerging song into reproducible syllable units of various types. Here we present a complementary approach of partitioning song development vertically into sequences of vocal changes. Note that identifying units of vocal change requires a smooth transition from the primitive to the mature version of a song interval, but it does not require classification of sounds in the emerging song. The imitation trajectory described above is an example of a DVC, as it has a limited scope in both song time and developmental time: We can identify the boundaries of the change by tracing the drop in the amplitude envelope. To estimate the attenuation we divide the syllable into three equal parts, measure minimum and maximum amplitude for each part and define: Attenuation ð%þ Minimumamp ðmiddlepartþ ¼ 1 Average fmaxampðfirstpartþ; MaxampðlastpartÞg ; ð1þ which is simply the ratio between the depth of the drop (minimum amplitude of the middle part) and the mean maximal amplitudes of first and last parts. We can now estimate the rate of the change by measuring the depth of the drop from day to day (Fig. 2C) and from hour to hour (Fig. 2D). As shown, the attenuation progressed slowly during the afternoon of day 52 post-hatch and rapidly during the morning of day 53. In addition to the limited scope, there are two features that add interest and generality to this vocal change: First, this was not an idiosyncratic change, but a step in a learning process, as it eventually led to an approximation of model sounds. Second, the acoustic feature that has changed (amplitude) can be studied in specific mechanistic terms: attenuation can be achieved, for example, by reducing air pressure or by briefly obstructing the upper airways (similar to inserting a stop consonant in human speech). Developmentally diachronic constraints The example discussed above is of an isolated vocal change. However, the song-imitation process involves

4 870 many vocal changes, some of them related. The imitation trajectory we have just examined (Fig. 2B D) is in fact more complex than the description given above suggests. As we described, the bird inserted a stop, which generated two indistinguishable harmonic stacks. Subsequently, the bird started to increase the amplitude of the second sound until it became about 50% louder than the first sound, so that a continuous prototype was
5 871 b Fig. 2 A an illustration of a song interval composed of different sounds, which can be automatically aligned during an extended developmental time as long as vocal changes are gradual. The red rectangle shows the boundaries of a distinctive vocal change (DVC). Note that in practice, the interval of sound is analyzed continuously and the symbols stand for aligned FFT time windows of sound rather than syllables. B An automatically traced imitation trajectory leading to a copy of the model harmonic stacks denoted as c and d. The bird inserted a stop, splitting a prototype harmonic stack into two sounds. C A quantitative tracing of the insertion of stop as it progresses from one day to the next. The blue curve represents the change in the depth of the attenuation (see blue amplitude curve in the spectral image). Each data point represents the mean value for 12 renditions of the syllable and the error bars represent the standard error (SE). D A quantitative tracing of the insertion of stop as it progresses from one hour to the next during the rapid phase of the vocal change (days 52 and 53 post-hatch). Each data point represents the mean value for 12 renditions of the syllable and the error bars represent SE eventually transformed into two distinguishable sounds. Figure 3A presents a 3-D image of the change in amplitude envelope during those vocal changes, suggesting that the amplitude of the second harmonic stack started increasing only after the stop was completed. To examine the dynamics of this amplitude modulation we use the same measurements as in Eq. 1, namely dividing the syllable into three equal parts, measuring minimum and maximum amplitude for each part and defining: relative amplitude ¼ Max amp ðlast partþ Max amp ðfirst partþ ; ð2þ which gives a value of 1 when there is no amplitude difference, and more than 1 when the second sound is louder than the first. As shown in Fig. 3B, the two vocal changes had very different time-courses: the insertion of stop occurred within a few days, whereas the amplification of the second sound progressed slowly over 20 days or so. Furthermore, the data suggest that the onset of amplification occurs only after the stop was inserted: For example, on day 53, 10:00 a.m. (Fig. 2D) the mean attenuation was 69 81% (95% confidences interval, n=12 renditions) indicating a statistically significant attenuation, and the mean relative amplitude was (95% confidence interval, n=12 renditions) which does not suggest a real amplitude modulation. To further validate that the onset of amplification occurs only after the stop was inserted we plot for each syllable the attenuation (Eq. 1) versus the amplification (relative amplitude, Eq. 2). Figure 3C presents the plot for all data sampled during the vocal change (on days 50 60). As shown, relative amplitude values are distributed evenly around 1 until attenuation levels are above 80% (which is virtually a stop). In other words, a tendency to increase the relative amplitude of the second part is detectable only in syllables where the insertion of stop is at a very advanced stage or complete. It seems likely that the outcome of the first vocal change is relevant (and perhaps even necessary) for activating the next vocal change. When a vocal change appears to be contingent on the completion of a different vocal change, we call it a developmentally diachronic constraint. Such constraints are manifested in the imitation trajectory as a function of developmental time (the y-axis of Fig. 4A). For example, when one vocal change (blue rectangle, c, dfic, D) produces the input of the next vocal change (red trapezoid, DfiD ), we say that the first vocal change is a developmentally diachronic constraint on the second one (Fig. 4A). Overall, song becomes more structured during development and indeed, each of the vocal changes described above (insertion of stop and amplitude modulation) added some structure to the song: the first generated two sounds out of one, and the second vocal change made them distinguishable. The second vocal change, however, did not approximate the model: in the song model, the amplitude of the first harmonic stack was higher than that of the second and hence, by amplifying the second harmonic stack the bird reduced the similarity to the song model. A similar phenomenon is also apparent in the next example: The correction of pitch error via the period-doubling trajectory presented in Fig. 3D, E is another example of developmentally diachronic constraint (from Tchernichovski et al. 2001). In this case, the first vocal change gradually increased the pitch, drifting it away from that of the model sound until it was twice the pitch of the model sound. At that point, a second vocal change halved the pitch in a single step (period doubling), matching it to that of the model. Here also, the gradual increase of pitch is likely to be a developmentally diachronic constraint required for the expression of period doubling. If the period doubling event must wait for the conclusion of the pitch increase, it might be possible to prevent the period doubling by delaying the learning. For example, after the age of 90 days song structure does not change much, if at all. The question is: had the first vocal change (the gradual increase of pitch) failed to reach the first harmonic of the song model by day 90, would the pitch freeze on that level? This has not been investigated yet, but we have seen a few cases of slowlearning birds in which period doubling failed to occur, and the pitch remained almost twice as high as that of the model. Developmentally synchronic constraints The challenge of song imitation is not only of changing sounds appropriately, but also of incorporating those changes into the emerging song structure. Consider, for example, a vocal change that expands the duration of a sound: DfiD (red trapezoid, Fig. 4A). This vocal change could be constrained by the need to shift the neighboring sounds (and of increasing the duration of the song motif). Constraints on vocal changes in song time (x-axis) are defined as developmentally synchronic constraints, as opposed to developmentally diachronic
: Note that the model song is shown twice, aligned with the start of the pupil s song on day 50 (bottom of figure) and with the end of the pupil s song on day 95 (top of figure).")
6 872 constraints we discussed earlier, which act in developmental time (y-axis). We will illustrate this by presenting an interesting case (Fig. 4B): Note that the model song is shown twice, aligned with the start of the pupil s song on day 50 (bottom of figure) and with the end of the pupil s song on day 95 (top of figure). As indicated by the arrows, by day 50 post-hatch a bird had already copied a raw version of the beginning and end of the song model but has not yet copied the harmonic stacks c. In order to copy
7 873 b Fig. 3 A A 3-D image of the change in the amplitude envelope during and after inserting a stop. To eliminate the effect of proximity to the microphone, we normalized each amplitude curve by the mean amplitude during the first 100 ms of the sound. As shown, the relative amplitude of the second call increases after the stop until it became 50% higher than that of the first call. B A quantitative tracing of the amplitude modulation (red line) in reference to the insertion of stop (blue line, both curves are 5th order polynominal smoothing). C A plot of attenuation (insertion of stop) versus amplitude modulation for each syllable sampled throughout the period of vocal change (days 50 60). D, E Perioddoubling trajectory: the pitch error (the difference between bird s pitch and model pitch) increases smoothly until it reach the first harmonic of the song model, and is then corrected abruptly by period doubling (from Tchernichovski et al. 2001) sound c in its proper temporal context, the bird must open a gap (insert a time slot) and generate an additional sound in the middle of its emerging song; otherwise, it would overwrite an existing match. This requirement is imposed by the song structure present on day 50, and is therefore a developmentally synchronic constraint on imitating that harmonic stack: As shown in Fig. 4B, between days 50 and 95, the bird stretched the duration of the chirp denoted as b, and transformed it to the harmonic stack denoted as c (bfic). That is, the bird has transformed a copy of model-sound b to a copy of model-sound c. Note that the alignment of the sound with the song model had changed while it was transformed, as indicated by its alignment in reference to the end of the song model (top of figure) as opposed to its alignment in reference to the beginning of the song model in the earlier version (bottom of figure). During the transformation, the sound was not only stretched (time warped) but was also separated from the syllable just prior to it, as all the sounds to the left of the vocal change were shifted farther to the left, opening a 50-ms time slot that gave room for the time warping. Thus, in order to transform bfic the bird had shifted neighboring sounds, overcoming a developmentally synchronic constraint on the time warping. With regards to overwriting existing sounds, the interpretation is not straightforward, but we would like to make two comments. The location of a mismatch between the emerging song (on day 50) and the song model depends on how they are aligned: Aligning from the beginning gives good match between the two songs until we reach sound e. Aligning from the end, however, gives a good match until we reach model sound c. Therefore, overwriting c (as opposed to e), suggests that the bird detected a mismatch by aligning its song in reference to the end of the song model. If this hypothesis is true, than the bird could not have detected the mismatch in real time (Margoliash 2002). Second, it is not clear if the overwriting reflects a failure to insert a sufficiently prolonged time slot, a failure to generate an additional sound, or a failure to detect that it is overwriting. Note that the order of mastering sounds (in developmental time) can determine the nature of developmentally synchronic constraints in song time. For example, if instead of copying the beginning and the end of the song model first and then attempting to copy the middle (ACfiABC) the bird had copied the beginning and the middle first and then attempted to copy the end (ABfiABC) there would have been no need for opening a time slot and for shifting sounds. The sensitivity of developmentally synchronic constraints to the order of copying model sounds is the subject of the next sections. Analysis of imitation trajectories across birds We have seen in one bird (Fig. 4B) that changing the position of sounds in song time might be subject to developmentally synchronic constraints. What developmental factors could dictate that an initial position of a sound differs from its target position in song time? Figure 5A presents a case of a bird (pupil 1) that copied all the model sounds in good approximation to their position in the song model, whereas Fig. 5B shows another case (pupil 5) where a copy of model sound a (vibrato, red arrow) appears in a position that differs from that of the model. Why is the position of the vibrato correct in one bird and incorrect in another bird? Tracing imitation trajectories of model sounds d and e showed similarities in trajectories of 10 out of 12 birds that were trained with the same song model (Tchernichovski et al. 2001). Figure 5C presents five examples of such trajectories. As shown, the prototype sounds (broadband downsweeps) are similar across birds. In each case, a broadband noisy sound was transformed into a copy of the model harmonic stack d. The fate of the high-pitch sounds, however, varied across birds: pupil 3 deleted the first high pitched sound very late (red arrow). Pupils 1 and 2 deleted the first high-pitched sound earlier, and pupils 4 and 5 did not delete the first high-pitch sound at all, but transformed it into a vibrato. Note that the final copy of sounds d and e in birds that retained the first high pitched sound (pupils 4 and 5) are similar to immature versions of those sounds in birds that deleted that high-pitched sound (e.g., compare to pupil 2, day 55). We propose that retaining this high-pitch sound and transforming it into a vibrato imposes developmentally synchronic constraints that cannot be fully resolved, and which can explain why pupils 1, 2 and 3 chose to delete that high-pitch prototype. In other words, we suggest that the decision of how to change one sound imposes constraints on the imitation of other sounds later on, and that taking those constraints into account may help the bird achieve a good imitation. Statistical analysis of imitation priorities The cases presented in Fig. 5C suggest that imitation trajectories of different birds share common features. As we suggested, similarity in vocal imitation across birds could stem from developmental constraints, but it could also stem from (or be modulated by) perceptual biases,
.")
8 874 Fig. 4 A An illustration of developmental synchronic constraints (horizontal arrow) versus developmentally diachronic constraints (vertical arrow) on a vocal changes. B Examining the emerging song motif of a bird on day 50 show imitation of the beginning and end of the song model (black arrows). The imitation trajectory between days 50 and 95 show that a copy of the chirp denoted by b was transformed into a copy of the harmonic stack c or priorities (ten Cate 1994) to copy certain speciesspecific sounds. This idea was introduced by Marler and Nelson (1992) to explain why distant populations of birds retain species-specific characteristics in their songs despite the enormous range of different sounds that they can master. These two issues are related, as within the context of copying a specific song model, a tendency to copy some sounds before others may impose developmentally synchronic constraints on the learning process. In other words, when a bird copies sound A and then sound B, it could be either because it prefers sound A or because it is easier for the bird to copy A before B. Note that the first explanation is purely perceptual: the bird gives priority to a subset of the song model acoustic structure. The second explanation has an ultimate motor cause, which comes about as primitive sounds are altered as the bird strives to match the model. We will now

9 875 Fig. 5 A The mature song of pupil 1 compared to the model song. B The mature song of pupil 5 compared to the model song. C Automatically traced imitation trajectories of sounds d and e in five birds (pupils 1 and 5 are the same as those in Fig. 5A and B, respectively) examine if imitation priorities are indeed similar across birds and if those priorities imposes similar developmentally synchronic constraints. To uncover the priority of copying different parts of a song model we trained nine birds very briefly (for only 2 days, overall about 1 min of playbacks; see Materials and methods) so as to induce partial imitation of the model song. In particular, we wanted to examine if the brief training would cause impairment specific to one part of the model song. The results of brief training are judged in reference to those of moderate training with the same song model: As shown in Fig. 6A, brief training was sufficient to induce some imitation, with the similarity to the last part of the song model being high across birds and the similarity to the middle part of the
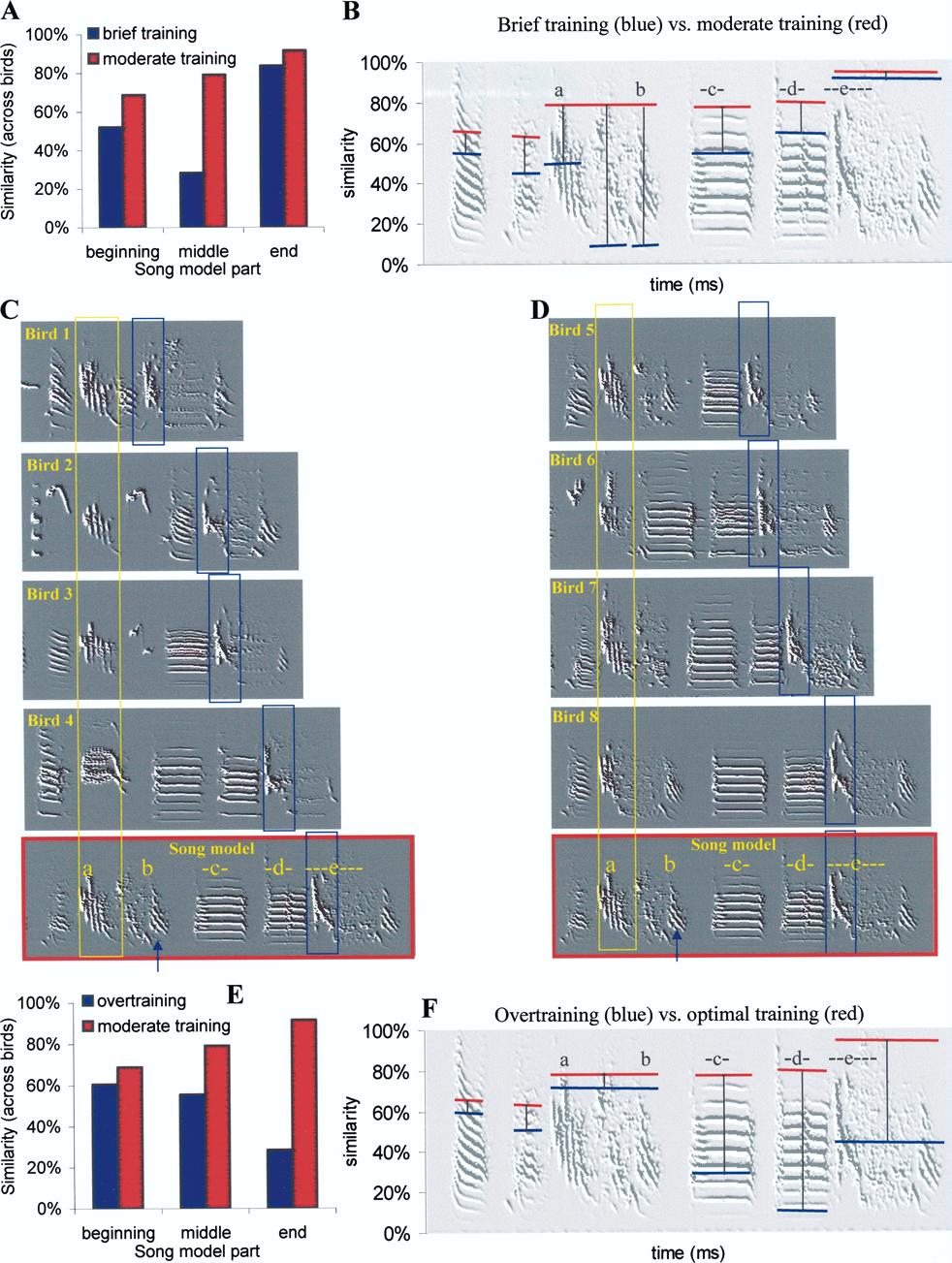
10 876
11 877 b Fig. 6 A We divided the song model to three equal parts (beginning, middle, and end) and calculated the mean similarity (% significant imitation) of each part for songs of the brief training group (blue bars) and for songs of the moderate training group (red bars). B A breakdown of similarity for each syllable separately. The red lines represent the proportion of moderate training birds that copied each syllable and the blue lines represent the proportion of brief training birds that copied each syllable. Vertical bars represent the difference between groups. C, D Examples of mature song motifs in four birds that received brief training (C) versus four birds that received moderate training (D). The yellow rectangles point to copies of the model vibrato, whereas the blue rectangles point to copies of the glissando. Note that in all birds the duration between the two rectangles was shorter than that of the model song. E A comparison of similarity to beginning, middle, and end parts of the song model across moderate training (red bars) versus over training (blue bars) groups. F A breakdown of similarity for each syllable separately. The red lines represent the proportion of moderate training birds that copied each syllable and the blue lines represent the proportion of overtraining birds that copied each syllable model being low across birds. Comparing similarity values to first, middle, and last part of the model song across birds showed statistically significant differences across groups (Kruskal-Wallis ANOVA, H=6.8, P=0.034). This bias in the brief training group was particularly strong in sound b (Fig. 6B). Interestingly, this is the same sound that was overwritten by the bird shown in Fig. 4B. We will now examine a posteriori the variability in song structure within and across groups. Figure 6C presents four examples of songs of the brief training group where we could unequivocally identify copies of model sounds a (vibrato, blue rectangle) and e (glissando, yellow rectangle). We observed two effects: first, the time between sounds a and e was much shorter in the pupil s songs than in the song model (184±66 ms in birds of the brief training group versus 405 ms in the song model, mean±sd throughout). Second, the sounds in the neighborhood of sound a (vibrato) varied greatly across birds (similarity values across birds in a 100-ms interval around the vibrato was only 9±14%), whereas the sounds in the neighborhood of the glissando (near the boundaries of the blue rectangle) were much more similar across birds (63±19%). Interestingly, similar effects (albeit weaker) were observed also in birds that received moderate training. As shown in Fig. 6D, the time between the two landmarks in birds of the moderate training group was shorter than that of the song model (290±84 ms versus 405 ms in the song model, judged in five out of six birds of this group where copies of sounds a and e were identified). The sounds in the neighborhood of the vibrato were variable across birds (33±32% similarity), whereas the sounds in the neighborhood of the glissando were very similar across birds (70±29% similarity). Although we did not record song development in the brief training and moderate training groups presented above, the results are consistent with those presented in Fig. 5, i.e., we observed a tendency to copy sound a (vibrato) in a position which is too close to that of sound e. This tendency, however, was strong in birds that received brief training, and weak in birds that received moderate training between days 30 and 90 post-hatch, where imitation is most accurate. Therefore, the priority of briefly trained birds to copy the beginning and end of the song model could be a consequence of different choices made during early stages of song development, as illustrated in Fig. 5C. The results of the overtrained groups were very different from those documented above: although the overtrained birds were exposed to the same song model during the same developmental time, they showed different imitation priorities. As shown in Fig. 6E, imitation of the last part of the song model was less accurate (similarity value of 23%) in the overtrained birds, whereas the first and middle parts of the song model were copied much more accurately. Note that this result is very different from that shown in Fig. 6A. As shown in Fig. 6F, the vibrato and the neighboring sounds were copied accurately, whereas most birds did not copy the harmonic stacks and the glissando. Since we did not record song development in these birds we do not know what constraints could have shaped their song development. Nevertheless, it suggests that imitation priorities are not fully determined by the acoustic structure of the model sounds. Discussion The purpose of this investigation was to suggest an approach for identifying units of vocal change and to uncover constraints that govern their expression during song imitation in the zebra finch. So far we have only examined a few simple cases and we do not know how much of the vocal learning process could be captured by the approach described here. Nevertheless, documenting the variety of DVCs across birds and across song models can would the toolbox of vocal changes available to the bird for achieving vocal learning. Since vocal imitation is a oneway process, it would be interesting to search for symmetry across vocal changes: for example, can a bird accomplish the inverse of inserting a stop, i.e., merging two sounds with equal facility? It will also be useful to examine what DVCs are available during different developmental stages. Finally, it would be interesting to examine individual variability in the toolbox of available vocal changes, e.g., across good and poor learners. Inherited phenotypes of birds that differ in vocal learning have been identified in Canaries (Mundinger 1995) and it would be useful to have similar preparations in the zebra finch, where both sound analysis and brain measurements are easier to perform. We suggest that some understanding of the vocal learning procedure and its underlying mechanisms can be achieved by careful analysis of developmentally synchronic and developmentally diachronic constraints on vocal changes. In particular, identifying
12 878 conservative and tightly coupled sequences of vocal changes leading to imitation would provide insight into the nature of the computational steps involved in sensory motor transformation. For example, period-doubling trajectories have the appearance of a two-stage approximation mechanism. A systematic analysis of many vocal changes could perhaps tell us, more generally, how many steps are involved in approximating specific sounds, and what level of control system is at play (Powers 1973). Units of vocal change and constraints on their expression can be explored across behavioral, perceptual (ten Cate 1994), articulatory (Suthers 1999; Goller and Larsen 2002; Fee 2002; Vicario 1991) and central (Jarvis et al. 2002; Lucas et al. 2002; Solis and Doupe 1999; Yu and Margoliash 1996) levels. For example, relating vocal changes to specific articulatory gestures could potentially generalize the descriptive model since the same vocal change could be triggered by more than one gesture and the same gesture could trigger more than one vocal change (Fee et al. 1998) or trigger different vocal changes in different sounds. Similar relations might exist between central and peripheral dynamics (Bernstein 1967; Brezina et al. 2000) of song learning. Overall, the song system provides a unique opportunity to study the information flow across system levels from moment to moment, under tight experimental control, and detailed behavioral analysis of song development would be necessary for understanding vocal learning mechanisms across those levels. Acknowledgements We thank Thierry Lints and Joshua Wallman for useful comments. Supported by the US Public Health Service (PHS) grant DC and by NIH RCMI grant GR12RR to CCNY. The experiments comply with the Principles of Animal Care, publication No , revised 1985, of the National Institutes of Health and also with the current laws of the USA. References Adret P (1993) Operant conditioning, song learning and imprinting to taped song in the zebra finch. Anim Behav 46: Bernstein N (1967) The co-ordination and regulation of movement. Pergamon Press, London Brainard MS, Doupe AJ (2000) Interruption of a basal gangliaforebrain circuit prevents plasticity of learned vocalizations. Nature 404:6779 Brezina V, Orekhova IV, Weiss K (2000) The neuromuscular transform: the dynamic, nonlinear link between motor neuron firing patterns and muscle contraction in rhythmic behaviors. J Neurophysiol 83: Cate C ten (1994) Perceptual mechanisms in imprinting and song learning. In: Hogan JA, Bolhuis JJ (eds) Causal mechanisms of behavioural development. Cambridge University Press, Cambridge, UK, pp Clark CW, Marler P, Beeman K (1987) Quantitative analysis of animal vocal phonology: an application to swamp sparrow song. Ethology 76:101 Goller F, Larsen ON (2002) New perspectives on mechanisms of sound generation in songbirds. J Comp Physiol (in press). DOI /s Fee MS (2002) Measurement of the linear and nonlinear mechanical properties of the oscine syrinx: implications for function. J Comp Physiol (in press). DOI /s z Fee MS, Shraiman B, Pesaran B, Mitra PP (1998) The role of nonlinear dynamics of the syrinx in the vocalizations of the songbird. Nature 395:67 Immelmann K (1969) Song development in the zebra finch and in other estrildid finches. In: Hinde R (ed) Bird vocalization. Cambridge University Press, Cambridge, pp Jarvis ED, Smith VA, Rivas1 MV, Wada K, McElroy M, Smulders TV, Carnici P, Hayashisaki Y, Dietrich F, Wu X, McConnell P, Wang P, Lin S (2002) Integrating the songbird brain. J Comp Physiol (in press) Konishi M (1965) The role of auditory feedback in the control of vocalization in the white-crowned sparrow. Z Tierpsychol 22:770 Lucas JR, Freeberg TM, Krishnan A, Long GR (2002) A comparative study of avian auditory brainstem responses: correlations with phylogeny and vocal complexity, and seasonal effects. J Comp Physiol (in press). DOI /s x Marler P, Nelson D (1992) Neuroselection and song learning in birds: species universals in culturally transmitted behavior. In: Marler P (ed) Seminars on the neurosciences, vol 4. Communication: behavior and neurobiology. Saunders, London, pp Margoliash D (2002) Evaluating theories of bird song learning: implications for future directions. J Comp Physiol (in press) Mundinger PC (1995) Behaviour-genetic analysis of canary song: inter-strain differences in sensory learning, and epigenetic rules. Anim Behav 50: Powers TP (1973) Behavior: the control of perception. Aldine de Gruyter Press, New York Price P (1979) Developmental determinants of structure in zebra finch song. J Comp Physiol Psychol 93:260 Solis MM, Doupe AJ (1999) Contributions of tutor and bird s own song experience to neural selectivity in the songbird anterior forebrain. J Neurosci 19: Suthers R (1999) The motor basis for vocal performance in songbirds. In: Hauser M, Konishi M (eds) The design of animal communication. MIT Press, Cambridge MA Tchernichovski O, Mitra PP (2002) Sound Analysis 2 software, user manual and song development database. Source: ofer.sci.ccny.cuny.edu Tchernichovski O, Lints T, Mitra PP, Nottebohm F (1999) Vocal imitation in zebra finches is inversely related to model abundance. Proc Natl Acad Sci. USA 96:12901 Tchernichovski O, Nottebohm F, Ho CE, Bijan P, Mitra PP (2000) A procedure for an automated measurement of song similarity. Anim Behav 59: Tchernichovski O, Mitra, PP, Lints T, Nottebohm F (2001) Dynamics of the vocal imitation process: how a zebra finch learns its song. Science 291: Thorpe WH (1958) The learning of song patterns by birds, with especial reference to the song of the chaffinch, Fringella coelebs. Ibis 100:535 Yu AC, Margoliash D (1996) Temporal hierarchical control of singing in birds. Science 273: Vicario DS (1991) Contribution of syringeal muscles to respiration and vocalization in the zebra finch. J Neurobiol 22:1
A Technique for Characterizing the Development of Rhythms in Bird Song
 A Technique for Characterizing the Development of Rhythms in Bird Song Sigal Saar 1,2 *, Partha P. Mitra 2 1 Department of Biology, The City College of New York, City University of New York, New York,
A Technique for Characterizing the Development of Rhythms in Bird Song Sigal Saar 1,2 *, Partha P. Mitra 2 1 Department of Biology, The City College of New York, City University of New York, New York,
Olga Feher, PhD Dissertation: Chapter 4 (May 2009) Chapter 4. Cumulative cultural evolution in an isolated colony
 Chapter 4. Cumulative cultural evolution in an isolated colony") Chapter 4. Cumulative cultural evolution in an isolated colony Background & Rationale The first time the question of multigenerational progression towards WT surfaced, we set out to answer it by recreating
Chapter 4. Cumulative cultural evolution in an isolated colony Background & Rationale The first time the question of multigenerational progression towards WT surfaced, we set out to answer it by recreating
A procedure for an automated measurement of song similarity
 ANIMAL BEHAVIOUR, 2000, 59, 000 000 doi:10.1006/anbe.1999.1416, available online at http://www.idealibrary.com on A procedure for an automated measurement of song similarity OFER TCHERNICHOVSKI*, FERNANDO
ANIMAL BEHAVIOUR, 2000, 59, 000 000 doi:10.1006/anbe.1999.1416, available online at http://www.idealibrary.com on A procedure for an automated measurement of song similarity OFER TCHERNICHOVSKI*, FERNANDO
DAT335 Music Perception and Cognition Cogswell Polytechnical College Spring Week 6 Class Notes
 DAT335 Music Perception and Cognition Cogswell Polytechnical College Spring 2009 Week 6 Class Notes Pitch Perception Introduction Pitch may be described as that attribute of auditory sensation in terms
DAT335 Music Perception and Cognition Cogswell Polytechnical College Spring 2009 Week 6 Class Notes Pitch Perception Introduction Pitch may be described as that attribute of auditory sensation in terms
Measurement of overtone frequencies of a toy piano and perception of its pitch
 Measurement of overtone frequencies of a toy piano and perception of its pitch PACS: 43.75.Mn ABSTRACT Akira Nishimura Department of Media and Cultural Studies, Tokyo University of Information Sciences,
Measurement of overtone frequencies of a toy piano and perception of its pitch PACS: 43.75.Mn ABSTRACT Akira Nishimura Department of Media and Cultural Studies, Tokyo University of Information Sciences,
2. AN INTROSPECTION OF THE MORPHING PROCESS
 1. INTRODUCTION Voice morphing means the transition of one speech signal into another. Like image morphing, speech morphing aims to preserve the shared characteristics of the starting and final signals,
1. INTRODUCTION Voice morphing means the transition of one speech signal into another. Like image morphing, speech morphing aims to preserve the shared characteristics of the starting and final signals,
Behavioral and neural identification of birdsong under several masking conditions
 Behavioral and neural identification of birdsong under several masking conditions Barbara G. Shinn-Cunningham 1, Virginia Best 1, Micheal L. Dent 2, Frederick J. Gallun 1, Elizabeth M. McClaine 2, Rajiv
Behavioral and neural identification of birdsong under several masking conditions Barbara G. Shinn-Cunningham 1, Virginia Best 1, Micheal L. Dent 2, Frederick J. Gallun 1, Elizabeth M. McClaine 2, Rajiv
Acoustic and neural bases for innate recognition of song
 Proc. Natl. Acad. Sci. USA Vol. 94, pp. 12694 12698, November 1997 Neurobiology Acoustic and neural bases for innate recognition of song C. S. WHALING*, M. M. SOLIS, A.J.DOUPE, J.A.SOHA*, AND P. MARLER*
Proc. Natl. Acad. Sci. USA Vol. 94, pp. 12694 12698, November 1997 Neurobiology Acoustic and neural bases for innate recognition of song C. S. WHALING*, M. M. SOLIS, A.J.DOUPE, J.A.SOHA*, AND P. MARLER*
Atypical Song Reveals Spontaneously Developing Coordination between Multi-Modal Signals in Brown- Headed Cowbirds (Molothrus ater)
") Atypical Song Reveals Spontaneously Developing Coordination between Multi-Modal Signals in Brown- Headed Cowbirds (Molothrus ater) Amanda R. Hoepfner*, Franz Goller Department of Biology, University of
Atypical Song Reveals Spontaneously Developing Coordination between Multi-Modal Signals in Brown- Headed Cowbirds (Molothrus ater) Amanda R. Hoepfner*, Franz Goller Department of Biology, University of
Computer Coordination With Popular Music: A New Research Agenda 1
 Computer Coordination With Popular Music: A New Research Agenda 1 Roger B. Dannenberg roger.dannenberg@cs.cmu.edu http://www.cs.cmu.edu/~rbd School of Computer Science Carnegie Mellon University Pittsburgh,
Computer Coordination With Popular Music: A New Research Agenda 1 Roger B. Dannenberg roger.dannenberg@cs.cmu.edu http://www.cs.cmu.edu/~rbd School of Computer Science Carnegie Mellon University Pittsburgh,
Chapter 4 : An efference copy may be used to maintain the stability of adult birdsong
 LMAN dynamics underlying normal and perturbed singing 45 Chapter 4 : An efference copy may be used to maintain the stability of adult birdsong Zebra finches use auditory feedback to both learn and maintain
LMAN dynamics underlying normal and perturbed singing 45 Chapter 4 : An efference copy may be used to maintain the stability of adult birdsong Zebra finches use auditory feedback to both learn and maintain
Estimating the Time to Reach a Target Frequency in Singing
 THE NEUROSCIENCES AND MUSIC III: DISORDERS AND PLASTICITY Estimating the Time to Reach a Target Frequency in Singing Sean Hutchins a and David Campbell b a Department of Psychology, McGill University,
THE NEUROSCIENCES AND MUSIC III: DISORDERS AND PLASTICITY Estimating the Time to Reach a Target Frequency in Singing Sean Hutchins a and David Campbell b a Department of Psychology, McGill University,
PS User Guide Series Seismic-Data Display
 PS User Guide Series 2015 Seismic-Data Display Prepared By Choon B. Park, Ph.D. January 2015 Table of Contents Page 1. File 2 2. Data 2 2.1 Resample 3 3. Edit 4 3.1 Export Data 4 3.2 Cut/Append Records
PS User Guide Series 2015 Seismic-Data Display Prepared By Choon B. Park, Ph.D. January 2015 Table of Contents Page 1. File 2 2. Data 2 2.1 Resample 3 3. Edit 4 3.1 Export Data 4 3.2 Cut/Append Records
Pitch. The perceptual correlate of frequency: the perceptual dimension along which sounds can be ordered from low to high.
 Pitch The perceptual correlate of frequency: the perceptual dimension along which sounds can be ordered from low to high. 1 The bottom line Pitch perception involves the integration of spectral (place)
Pitch The perceptual correlate of frequency: the perceptual dimension along which sounds can be ordered from low to high. 1 The bottom line Pitch perception involves the integration of spectral (place)
Using the new psychoacoustic tonality analyses Tonality (Hearing Model) 1
 1") 02/18 Using the new psychoacoustic tonality analyses 1 As of ArtemiS SUITE 9.2, a very important new fully psychoacoustic approach to the measurement of tonalities is now available., based on the Hearing
02/18 Using the new psychoacoustic tonality analyses 1 As of ArtemiS SUITE 9.2, a very important new fully psychoacoustic approach to the measurement of tonalities is now available., based on the Hearing
KNX Dimmer RGBW - User Manual
 KNX Dimmer RGBW - User Manual Item No.: LC-013-004 1. Product Description With the KNX Dimmer RGBW it is possible to control of RGBW, WW-CW LED or 4 independent channels with integrated KNX BCU. Simple
KNX Dimmer RGBW - User Manual Item No.: LC-013-004 1. Product Description With the KNX Dimmer RGBW it is possible to control of RGBW, WW-CW LED or 4 independent channels with integrated KNX BCU. Simple
AUD 6306 Speech Science
 AUD 3 Speech Science Dr. Peter Assmann Spring semester 2 Role of Pitch Information Pitch contour is the primary cue for tone recognition Tonal languages rely on pitch level and differences to convey lexical
AUD 3 Speech Science Dr. Peter Assmann Spring semester 2 Role of Pitch Information Pitch contour is the primary cue for tone recognition Tonal languages rely on pitch level and differences to convey lexical
AN ARTISTIC TECHNIQUE FOR AUDIO-TO-VIDEO TRANSLATION ON A MUSIC PERCEPTION STUDY
 AN ARTISTIC TECHNIQUE FOR AUDIO-TO-VIDEO TRANSLATION ON A MUSIC PERCEPTION STUDY Eugene Mikyung Kim Department of Music Technology, Korea National University of Arts eugene@u.northwestern.edu ABSTRACT
AN ARTISTIC TECHNIQUE FOR AUDIO-TO-VIDEO TRANSLATION ON A MUSIC PERCEPTION STUDY Eugene Mikyung Kim Department of Music Technology, Korea National University of Arts eugene@u.northwestern.edu ABSTRACT
VOCAL TRACT FUNCTION IN BIRDSONG PRODUCTION: EXPERIMENTAL MANIPULATION OF BEAK MOVEMENTS
 The Journal of Experimental Biology 23, 1845 1855 (2) Printed in Great Britain The Company of Biologists Limited 2 JEB2489 1845 VOCAL TRACT FUNCTION IN BIRDSONG PRODUCTION: EXPERIMENTAL MANIPULATION OF
The Journal of Experimental Biology 23, 1845 1855 (2) Printed in Great Britain The Company of Biologists Limited 2 JEB2489 1845 VOCAL TRACT FUNCTION IN BIRDSONG PRODUCTION: EXPERIMENTAL MANIPULATION OF
Instrument Recognition in Polyphonic Mixtures Using Spectral Envelopes
 Instrument Recognition in Polyphonic Mixtures Using Spectral Envelopes hello Jay Biernat Third author University of Rochester University of Rochester Affiliation3 words jbiernat@ur.rochester.edu author3@ismir.edu
Instrument Recognition in Polyphonic Mixtures Using Spectral Envelopes hello Jay Biernat Third author University of Rochester University of Rochester Affiliation3 words jbiernat@ur.rochester.edu author3@ismir.edu
Application Note AN-708 Vibration Measurements with the Vibration Synchronization Module
 Application Note AN-708 Vibration Measurements with the Vibration Synchronization Module Introduction The vibration module allows complete analysis of cyclical events using low-speed cameras. This is accomplished
Application Note AN-708 Vibration Measurements with the Vibration Synchronization Module Introduction The vibration module allows complete analysis of cyclical events using low-speed cameras. This is accomplished
Modeling memory for melodies
 Modeling memory for melodies Daniel Müllensiefen 1 and Christian Hennig 2 1 Musikwissenschaftliches Institut, Universität Hamburg, 20354 Hamburg, Germany 2 Department of Statistical Science, University
Modeling memory for melodies Daniel Müllensiefen 1 and Christian Hennig 2 1 Musikwissenschaftliches Institut, Universität Hamburg, 20354 Hamburg, Germany 2 Department of Statistical Science, University
Laboratory Assignment 3. Digital Music Synthesis: Beethoven s Fifth Symphony Using MATLAB
 Laboratory Assignment 3 Digital Music Synthesis: Beethoven s Fifth Symphony Using MATLAB PURPOSE In this laboratory assignment, you will use MATLAB to synthesize the audio tones that make up a well-known
Laboratory Assignment 3 Digital Music Synthesis: Beethoven s Fifth Symphony Using MATLAB PURPOSE In this laboratory assignment, you will use MATLAB to synthesize the audio tones that make up a well-known
Pitch Perception and Grouping. HST.723 Neural Coding and Perception of Sound
 Pitch Perception and Grouping HST.723 Neural Coding and Perception of Sound Pitch Perception. I. Pure Tones The pitch of a pure tone is strongly related to the tone s frequency, although there are small
Pitch Perception and Grouping HST.723 Neural Coding and Perception of Sound Pitch Perception. I. Pure Tones The pitch of a pure tone is strongly related to the tone s frequency, although there are small
The Measurement Tools and What They Do
 2 The Measurement Tools The Measurement Tools and What They Do JITTERWIZARD The JitterWizard is a unique capability of the JitterPro package that performs the requisite scope setup chores while simplifying
2 The Measurement Tools The Measurement Tools and What They Do JITTERWIZARD The JitterWizard is a unique capability of the JitterPro package that performs the requisite scope setup chores while simplifying
THE DIGITAL DELAY ADVANTAGE A guide to using Digital Delays. Synchronize loudspeakers Eliminate comb filter distortion Align acoustic image.
 THE DIGITAL DELAY ADVANTAGE A guide to using Digital Delays Synchronize loudspeakers Eliminate comb filter distortion Align acoustic image Contents THE DIGITAL DELAY ADVANTAGE...1 - Why Digital Delays?...
THE DIGITAL DELAY ADVANTAGE A guide to using Digital Delays Synchronize loudspeakers Eliminate comb filter distortion Align acoustic image Contents THE DIGITAL DELAY ADVANTAGE...1 - Why Digital Delays?...
BER MEASUREMENT IN THE NOISY CHANNEL
 BER MEASUREMENT IN THE NOISY CHANNEL PREPARATION... 2 overview... 2 the basic system... 3 a more detailed description... 4 theoretical predictions... 5 EXPERIMENT... 6 the ERROR COUNTING UTILITIES module...
BER MEASUREMENT IN THE NOISY CHANNEL PREPARATION... 2 overview... 2 the basic system... 3 a more detailed description... 4 theoretical predictions... 5 EXPERIMENT... 6 the ERROR COUNTING UTILITIES module...
Object selectivity of local field potentials and spikes in the macaque inferior temporal cortex
 Object selectivity of local field potentials and spikes in the macaque inferior temporal cortex Gabriel Kreiman 1,2,3,4*#, Chou P. Hung 1,2,4*, Alexander Kraskov 5, Rodrigo Quian Quiroga 6, Tomaso Poggio
Object selectivity of local field potentials and spikes in the macaque inferior temporal cortex Gabriel Kreiman 1,2,3,4*#, Chou P. Hung 1,2,4*, Alexander Kraskov 5, Rodrigo Quian Quiroga 6, Tomaso Poggio
The Research of Controlling Loudness in the Timbre Subjective Perception Experiment of Sheng
 The Research of Controlling Loudness in the Timbre Subjective Perception Experiment of Sheng S. Zhu, P. Ji, W. Kuang and J. Yang Institute of Acoustics, CAS, O.21, Bei-Si-huan-Xi Road, 100190 Beijing,
The Research of Controlling Loudness in the Timbre Subjective Perception Experiment of Sheng S. Zhu, P. Ji, W. Kuang and J. Yang Institute of Acoustics, CAS, O.21, Bei-Si-huan-Xi Road, 100190 Beijing,
Nature Neuroscience: doi: /nn Supplementary Figure 1. Emergence of dmpfc and BLA 4-Hz oscillations during freezing behavior.
 Supplementary Figure 1 Emergence of dmpfc and BLA 4-Hz oscillations during freezing behavior. (a) Representative power spectrum of dmpfc LFPs recorded during Retrieval for freezing and no freezing periods.
Supplementary Figure 1 Emergence of dmpfc and BLA 4-Hz oscillations during freezing behavior. (a) Representative power spectrum of dmpfc LFPs recorded during Retrieval for freezing and no freezing periods.
Acoustic and musical foundations of the speech/song illusion
 Acoustic and musical foundations of the speech/song illusion Adam Tierney, *1 Aniruddh Patel #2, Mara Breen^3 * Department of Psychological Sciences, Birkbeck, University of London, United Kingdom # Department
Acoustic and musical foundations of the speech/song illusion Adam Tierney, *1 Aniruddh Patel #2, Mara Breen^3 * Department of Psychological Sciences, Birkbeck, University of London, United Kingdom # Department
Reducing tilt errors in moiré linear encoders using phase-modulated grating
 REVIEW OF SCIENTIFIC INSTRUMENTS VOLUME 71, NUMBER 6 JUNE 2000 Reducing tilt errors in moiré linear encoders using phase-modulated grating Ju-Ho Song Multimedia Division, LG Electronics, #379, Kasoo-dong,
REVIEW OF SCIENTIFIC INSTRUMENTS VOLUME 71, NUMBER 6 JUNE 2000 Reducing tilt errors in moiré linear encoders using phase-modulated grating Ju-Ho Song Multimedia Division, LG Electronics, #379, Kasoo-dong,
Musical Acoustics Lecture 15 Pitch & Frequency (Psycho-Acoustics)
") 1 Musical Acoustics Lecture 15 Pitch & Frequency (Psycho-Acoustics) Pitch Pitch is a subjective characteristic of sound Some listeners even assign pitch differently depending upon whether the sound was
1 Musical Acoustics Lecture 15 Pitch & Frequency (Psycho-Acoustics) Pitch Pitch is a subjective characteristic of sound Some listeners even assign pitch differently depending upon whether the sound was
OBJECTIVE EVALUATION OF A MELODY EXTRACTOR FOR NORTH INDIAN CLASSICAL VOCAL PERFORMANCES
 OBJECTIVE EVALUATION OF A MELODY EXTRACTOR FOR NORTH INDIAN CLASSICAL VOCAL PERFORMANCES Vishweshwara Rao and Preeti Rao Digital Audio Processing Lab, Electrical Engineering Department, IIT-Bombay, Powai,
OBJECTIVE EVALUATION OF A MELODY EXTRACTOR FOR NORTH INDIAN CLASSICAL VOCAL PERFORMANCES Vishweshwara Rao and Preeti Rao Digital Audio Processing Lab, Electrical Engineering Department, IIT-Bombay, Powai,
APPLICATIONS OF A SEMI-AUTOMATIC MELODY EXTRACTION INTERFACE FOR INDIAN MUSIC
 APPLICATIONS OF A SEMI-AUTOMATIC MELODY EXTRACTION INTERFACE FOR INDIAN MUSIC Vishweshwara Rao, Sachin Pant, Madhumita Bhaskar and Preeti Rao Department of Electrical Engineering, IIT Bombay {vishu, sachinp,
APPLICATIONS OF A SEMI-AUTOMATIC MELODY EXTRACTION INTERFACE FOR INDIAN MUSIC Vishweshwara Rao, Sachin Pant, Madhumita Bhaskar and Preeti Rao Department of Electrical Engineering, IIT Bombay {vishu, sachinp,
Proceedings of Meetings on Acoustics
 Proceedings of Meetings on Acoustics Volume 19, 2013 http://acousticalsociety.org/ ICA 2013 Montreal Montreal, Canada 2-7 June 2013 Psychological and Physiological Acoustics Session 1pPPb: Psychoacoustics
Proceedings of Meetings on Acoustics Volume 19, 2013 http://acousticalsociety.org/ ICA 2013 Montreal Montreal, Canada 2-7 June 2013 Psychological and Physiological Acoustics Session 1pPPb: Psychoacoustics
Real-time Granular Sampling Using the IRCAM Signal Processing Workstation. Cort Lippe IRCAM, 31 rue St-Merri, Paris, 75004, France
 Cort Lippe 1 Real-time Granular Sampling Using the IRCAM Signal Processing Workstation Cort Lippe IRCAM, 31 rue St-Merri, Paris, 75004, France Running Title: Real-time Granular Sampling [This copy of this
Cort Lippe 1 Real-time Granular Sampling Using the IRCAM Signal Processing Workstation Cort Lippe IRCAM, 31 rue St-Merri, Paris, 75004, France Running Title: Real-time Granular Sampling [This copy of this
CATHODE RAY OSCILLOSCOPE. Basic block diagrams Principle of operation Measurement of voltage, current and frequency
 CATHODE RAY OSCILLOSCOPE Basic block diagrams Principle of operation Measurement of voltage, current and frequency 103 INTRODUCTION: The cathode-ray oscilloscope (CRO) is a multipurpose display instrument
CATHODE RAY OSCILLOSCOPE Basic block diagrams Principle of operation Measurement of voltage, current and frequency 103 INTRODUCTION: The cathode-ray oscilloscope (CRO) is a multipurpose display instrument
Temporal coordination in string quartet performance
 International Symposium on Performance Science ISBN 978-2-9601378-0-4 The Author 2013, Published by the AEC All rights reserved Temporal coordination in string quartet performance Renee Timmers 1, Satoshi
International Symposium on Performance Science ISBN 978-2-9601378-0-4 The Author 2013, Published by the AEC All rights reserved Temporal coordination in string quartet performance Renee Timmers 1, Satoshi
Effects of Musical Training on Key and Harmony Perception
 THE NEUROSCIENCES AND MUSIC III DISORDERS AND PLASTICITY Effects of Musical Training on Key and Harmony Perception Kathleen A. Corrigall a and Laurel J. Trainor a,b a Department of Psychology, Neuroscience,
THE NEUROSCIENCES AND MUSIC III DISORDERS AND PLASTICITY Effects of Musical Training on Key and Harmony Perception Kathleen A. Corrigall a and Laurel J. Trainor a,b a Department of Psychology, Neuroscience,
Precise Digital Integration of Fast Analogue Signals using a 12-bit Oscilloscope
 EUROPEAN ORGANIZATION FOR NUCLEAR RESEARCH CERN BEAMS DEPARTMENT CERN-BE-2014-002 BI Precise Digital Integration of Fast Analogue Signals using a 12-bit Oscilloscope M. Gasior; M. Krupa CERN Geneva/CH
EUROPEAN ORGANIZATION FOR NUCLEAR RESEARCH CERN BEAMS DEPARTMENT CERN-BE-2014-002 BI Precise Digital Integration of Fast Analogue Signals using a 12-bit Oscilloscope M. Gasior; M. Krupa CERN Geneva/CH
Concert halls conveyors of musical expressions
 Communication Acoustics: Paper ICA216-465 Concert halls conveyors of musical expressions Tapio Lokki (a) (a) Aalto University, Dept. of Computer Science, Finland, tapio.lokki@aalto.fi Abstract: The first
Communication Acoustics: Paper ICA216-465 Concert halls conveyors of musical expressions Tapio Lokki (a) (a) Aalto University, Dept. of Computer Science, Finland, tapio.lokki@aalto.fi Abstract: The first
Toward a Computationally-Enhanced Acoustic Grand Piano
 Toward a Computationally-Enhanced Acoustic Grand Piano Andrew McPherson Electrical & Computer Engineering Drexel University 3141 Chestnut St. Philadelphia, PA 19104 USA apm@drexel.edu Youngmoo Kim Electrical
Toward a Computationally-Enhanced Acoustic Grand Piano Andrew McPherson Electrical & Computer Engineering Drexel University 3141 Chestnut St. Philadelphia, PA 19104 USA apm@drexel.edu Youngmoo Kim Electrical
1 Ver.mob Brief guide
 1 Ver.mob 14.02.2017 Brief guide 2 Contents Introduction... 3 Main features... 3 Hardware and software requirements... 3 The installation of the program... 3 Description of the main Windows of the program...
1 Ver.mob 14.02.2017 Brief guide 2 Contents Introduction... 3 Main features... 3 Hardware and software requirements... 3 The installation of the program... 3 Description of the main Windows of the program...
Music Radar: A Web-based Query by Humming System
 Music Radar: A Web-based Query by Humming System Lianjie Cao, Peng Hao, Chunmeng Zhou Computer Science Department, Purdue University, 305 N. University Street West Lafayette, IN 47907-2107 {cao62, pengh,
Music Radar: A Web-based Query by Humming System Lianjie Cao, Peng Hao, Chunmeng Zhou Computer Science Department, Purdue University, 305 N. University Street West Lafayette, IN 47907-2107 {cao62, pengh,
Experiment 13 Sampling and reconstruction
 Experiment 13 Sampling and reconstruction Preliminary discussion So far, the experiments in this manual have concentrated on communications systems that transmit analog signals. However, digital transmission
Experiment 13 Sampling and reconstruction Preliminary discussion So far, the experiments in this manual have concentrated on communications systems that transmit analog signals. However, digital transmission
MIE 402: WORKSHOP ON DATA ACQUISITION AND SIGNAL PROCESSING Spring 2003
 MIE 402: WORKSHOP ON DATA ACQUISITION AND SIGNAL PROCESSING Spring 2003 OBJECTIVE To become familiar with state-of-the-art digital data acquisition hardware and software. To explore common data acquisition
MIE 402: WORKSHOP ON DATA ACQUISITION AND SIGNAL PROCESSING Spring 2003 OBJECTIVE To become familiar with state-of-the-art digital data acquisition hardware and software. To explore common data acquisition
Spatial-frequency masking with briefly pulsed patterns
 Perception, 1978, volume 7, pages 161-166 Spatial-frequency masking with briefly pulsed patterns Gordon E Legge Department of Psychology, University of Minnesota, Minneapolis, Minnesota 55455, USA Michael
Perception, 1978, volume 7, pages 161-166 Spatial-frequency masking with briefly pulsed patterns Gordon E Legge Department of Psychology, University of Minnesota, Minneapolis, Minnesota 55455, USA Michael
On time: the influence of tempo, structure and style on the timing of grace notes in skilled musical performance
 RHYTHM IN MUSIC PERFORMANCE AND PERCEIVED STRUCTURE 1 On time: the influence of tempo, structure and style on the timing of grace notes in skilled musical performance W. Luke Windsor, Rinus Aarts, Peter
RHYTHM IN MUSIC PERFORMANCE AND PERCEIVED STRUCTURE 1 On time: the influence of tempo, structure and style on the timing of grace notes in skilled musical performance W. Luke Windsor, Rinus Aarts, Peter
SREV1 Sampling Guide. An Introduction to Impulse-response Sampling with the SREV1 Sampling Reverberator
 An Introduction to Impulse-response Sampling with the SREV Sampling Reverberator Contents Introduction.............................. 2 What is Sound Field Sampling?.....................................
An Introduction to Impulse-response Sampling with the SREV Sampling Reverberator Contents Introduction.............................. 2 What is Sound Field Sampling?.....................................
Analysis of local and global timing and pitch change in ordinary
 Alma Mater Studiorum University of Bologna, August -6 6 Analysis of local and global timing and pitch change in ordinary melodies Roger Watt Dept. of Psychology, University of Stirling, Scotland r.j.watt@stirling.ac.uk
Alma Mater Studiorum University of Bologna, August -6 6 Analysis of local and global timing and pitch change in ordinary melodies Roger Watt Dept. of Psychology, University of Stirling, Scotland r.j.watt@stirling.ac.uk
Figure 1: Media Contents- Dandelights (The convergence of nature and technology) creative design in a wide range of art forms, but the image quality h
 creative design in a wide range of art forms, but the image quality h") Received January 21, 2017; Accepted January 21, 2017 Lee, Joon Seo Sungkyunkwan University mildjoon@skku.edu Sul, Sang Hun Sungkyunkwan University sanghunsul@skku.edu Media Façade and the design identity
Received January 21, 2017; Accepted January 21, 2017 Lee, Joon Seo Sungkyunkwan University mildjoon@skku.edu Sul, Sang Hun Sungkyunkwan University sanghunsul@skku.edu Media Façade and the design identity
EFFECT OF REPETITION OF STANDARD AND COMPARISON TONES ON RECOGNITION MEMORY FOR PITCH '
 Journal oj Experimental Psychology 1972, Vol. 93, No. 1, 156-162 EFFECT OF REPETITION OF STANDARD AND COMPARISON TONES ON RECOGNITION MEMORY FOR PITCH ' DIANA DEUTSCH " Center for Human Information Processing,
Journal oj Experimental Psychology 1972, Vol. 93, No. 1, 156-162 EFFECT OF REPETITION OF STANDARD AND COMPARISON TONES ON RECOGNITION MEMORY FOR PITCH ' DIANA DEUTSCH " Center for Human Information Processing,
E X P E R I M E N T 1
 E X P E R I M E N T 1 Getting to Know Data Studio Produced by the Physics Staff at Collin College Copyright Collin College Physics Department. All Rights Reserved. University Physics, Exp 1: Getting to
E X P E R I M E N T 1 Getting to Know Data Studio Produced by the Physics Staff at Collin College Copyright Collin College Physics Department. All Rights Reserved. University Physics, Exp 1: Getting to
ADAPTATION TO DISPLACED AND DELAYED VISUAL FEEDBACK FROM THE HAND 1
 Journal ol Experimental Psychology 1966, Vol. 72, No. 6, 887-891 ADAPTATION TO DISPLACED AND DELAYED VISUAL FEEDBACK FROM THE HAND 1 RICHARD HELD, AGLAIA EFSTATHIOU, AND MARTHA GREENE Massachusetts Institute
Journal ol Experimental Psychology 1966, Vol. 72, No. 6, 887-891 ADAPTATION TO DISPLACED AND DELAYED VISUAL FEEDBACK FROM THE HAND 1 RICHARD HELD, AGLAIA EFSTATHIOU, AND MARTHA GREENE Massachusetts Institute
DELTA MODULATION AND DPCM CODING OF COLOR SIGNALS
 DELTA MODULATION AND DPCM CODING OF COLOR SIGNALS Item Type text; Proceedings Authors Habibi, A. Publisher International Foundation for Telemetering Journal International Telemetering Conference Proceedings
DELTA MODULATION AND DPCM CODING OF COLOR SIGNALS Item Type text; Proceedings Authors Habibi, A. Publisher International Foundation for Telemetering Journal International Telemetering Conference Proceedings
Skip Length and Inter-Starvation Distance as a Combined Metric to Assess the Quality of Transmitted Video
 Skip Length and Inter-Starvation Distance as a Combined Metric to Assess the Quality of Transmitted Video Mohamed Hassan, Taha Landolsi, Husameldin Mukhtar, and Tamer Shanableh College of Engineering American
Skip Length and Inter-Starvation Distance as a Combined Metric to Assess the Quality of Transmitted Video Mohamed Hassan, Taha Landolsi, Husameldin Mukhtar, and Tamer Shanableh College of Engineering American
Topic 10. Multi-pitch Analysis
 Topic 10 Multi-pitch Analysis What is pitch? Common elements of music are pitch, rhythm, dynamics, and the sonic qualities of timbre and texture. An auditory perceptual attribute in terms of which sounds
Topic 10 Multi-pitch Analysis What is pitch? Common elements of music are pitch, rhythm, dynamics, and the sonic qualities of timbre and texture. An auditory perceptual attribute in terms of which sounds
However, in studies of expressive timing, the aim is to investigate production rather than perception of timing, that is, independently of the listene
 Beat Extraction from Expressive Musical Performances Simon Dixon, Werner Goebl and Emilios Cambouropoulos Austrian Research Institute for Artificial Intelligence, Schottengasse 3, A-1010 Vienna, Austria.
Beat Extraction from Expressive Musical Performances Simon Dixon, Werner Goebl and Emilios Cambouropoulos Austrian Research Institute for Artificial Intelligence, Schottengasse 3, A-1010 Vienna, Austria.
The Future of EMC Test Laboratory Capabilities. White Paper
 The Future of EMC Test Laboratory Capabilities White Paper The complexity of modern day electronics is increasing the EMI compliance failure rate. The result is a need for better EMI diagnostic capabilities
The Future of EMC Test Laboratory Capabilities White Paper The complexity of modern day electronics is increasing the EMI compliance failure rate. The result is a need for better EMI diagnostic capabilities
Speech Recognition and Signal Processing for Broadcast News Transcription
 2.2.1 Speech Recognition and Signal Processing for Broadcast News Transcription Continued research and development of a broadcast news speech transcription system has been promoted. Universities and researchers
2.2.1 Speech Recognition and Signal Processing for Broadcast News Transcription Continued research and development of a broadcast news speech transcription system has been promoted. Universities and researchers
Simple Harmonic Motion: What is a Sound Spectrum?
 Simple Harmonic Motion: What is a Sound Spectrum? A sound spectrum displays the different frequencies present in a sound. Most sounds are made up of a complicated mixture of vibrations. (There is an introduction
Simple Harmonic Motion: What is a Sound Spectrum? A sound spectrum displays the different frequencies present in a sound. Most sounds are made up of a complicated mixture of vibrations. (There is an introduction
FLOW INDUCED NOISE REDUCTION TECHNIQUES FOR MICROPHONES IN LOW SPEED WIND TUNNELS
 SENSORS FOR RESEARCH & DEVELOPMENT WHITE PAPER #42 FLOW INDUCED NOISE REDUCTION TECHNIQUES FOR MICROPHONES IN LOW SPEED WIND TUNNELS Written By Dr. Andrew R. Barnard, INCE Bd. Cert., Assistant Professor
SENSORS FOR RESEARCH & DEVELOPMENT WHITE PAPER #42 FLOW INDUCED NOISE REDUCTION TECHNIQUES FOR MICROPHONES IN LOW SPEED WIND TUNNELS Written By Dr. Andrew R. Barnard, INCE Bd. Cert., Assistant Professor
THE EFFECT OF PERFORMANCE STAGES ON SUBWOOFER POLAR AND FREQUENCY RESPONSES
 THE EFFECT OF PERFORMANCE STAGES ON SUBWOOFER POLAR AND FREQUENCY RESPONSES AJ Hill Department of Electronics, Computing & Mathematics, University of Derby, UK J Paul Department of Electronics, Computing
THE EFFECT OF PERFORMANCE STAGES ON SUBWOOFER POLAR AND FREQUENCY RESPONSES AJ Hill Department of Electronics, Computing & Mathematics, University of Derby, UK J Paul Department of Electronics, Computing
An Empirical Analysis of Macroscopic Fundamental Diagrams for Sendai Road Networks
 Interdisciplinary Information Sciences Vol. 21, No. 1 (2015) 49 61 #Graduate School of Information Sciences, Tohoku University ISSN 1340-9050 print/1347-6157 online DOI 10.4036/iis.2015.49 An Empirical
Interdisciplinary Information Sciences Vol. 21, No. 1 (2015) 49 61 #Graduate School of Information Sciences, Tohoku University ISSN 1340-9050 print/1347-6157 online DOI 10.4036/iis.2015.49 An Empirical
Agilent PN Time-Capture Capabilities of the Agilent Series Vector Signal Analyzers Product Note
 Agilent PN 89400-10 Time-Capture Capabilities of the Agilent 89400 Series Vector Signal Analyzers Product Note Figure 1. Simplified block diagram showing basic signal flow in the Agilent 89400 Series VSAs
Agilent PN 89400-10 Time-Capture Capabilities of the Agilent 89400 Series Vector Signal Analyzers Product Note Figure 1. Simplified block diagram showing basic signal flow in the Agilent 89400 Series VSAs
Quarterly Progress and Status Report. Perception of just noticeable time displacement of a tone presented in a metrical sequence at different tempos
 Dept. for Speech, Music and Hearing Quarterly Progress and Status Report Perception of just noticeable time displacement of a tone presented in a metrical sequence at different tempos Friberg, A. and Sundberg,
Dept. for Speech, Music and Hearing Quarterly Progress and Status Report Perception of just noticeable time displacement of a tone presented in a metrical sequence at different tempos Friberg, A. and Sundberg,
Full Disclosure Monitoring
 Full Disclosure Monitoring Power Quality Application Note Full Disclosure monitoring is the ability to measure all aspects of power quality, on every voltage cycle, and record them in appropriate detail
Full Disclosure Monitoring Power Quality Application Note Full Disclosure monitoring is the ability to measure all aspects of power quality, on every voltage cycle, and record them in appropriate detail
Tempo and Beat Analysis
 Advanced Course Computer Science Music Processing Summer Term 2010 Meinard Müller, Peter Grosche Saarland University and MPI Informatik meinard@mpi-inf.mpg.de Tempo and Beat Analysis Musical Properties:
Advanced Course Computer Science Music Processing Summer Term 2010 Meinard Müller, Peter Grosche Saarland University and MPI Informatik meinard@mpi-inf.mpg.de Tempo and Beat Analysis Musical Properties:
Experiment 9A: Magnetism/The Oscilloscope
 Experiment 9A: Magnetism/The Oscilloscope (This lab s "write up" is integrated into the answer sheet. You don't need to attach a separate one.) Part I: Magnetism and Coils A. Obtain a neodymium magnet
Experiment 9A: Magnetism/The Oscilloscope (This lab s "write up" is integrated into the answer sheet. You don't need to attach a separate one.) Part I: Magnetism and Coils A. Obtain a neodymium magnet
PulseCounter Neutron & Gamma Spectrometry Software Manual
 PulseCounter Neutron & Gamma Spectrometry Software Manual MAXIMUS ENERGY CORPORATION Written by Dr. Max I. Fomitchev-Zamilov Web: maximus.energy TABLE OF CONTENTS 0. GENERAL INFORMATION 1. DEFAULT SCREEN
PulseCounter Neutron & Gamma Spectrometry Software Manual MAXIMUS ENERGY CORPORATION Written by Dr. Max I. Fomitchev-Zamilov Web: maximus.energy TABLE OF CONTENTS 0. GENERAL INFORMATION 1. DEFAULT SCREEN
Timbre blending of wind instruments: acoustics and perception
 Timbre blending of wind instruments: acoustics and perception Sven-Amin Lembke CIRMMT / Music Technology Schulich School of Music, McGill University sven-amin.lembke@mail.mcgill.ca ABSTRACT The acoustical
Timbre blending of wind instruments: acoustics and perception Sven-Amin Lembke CIRMMT / Music Technology Schulich School of Music, McGill University sven-amin.lembke@mail.mcgill.ca ABSTRACT The acoustical
The Relationship Between Auditory Imagery and Musical Synchronization Abilities in Musicians
 The Relationship Between Auditory Imagery and Musical Synchronization Abilities in Musicians Nadine Pecenka, *1 Peter E. Keller, *2 * Music Cognition and Action Group, Max Planck Institute for Human Cognitive
The Relationship Between Auditory Imagery and Musical Synchronization Abilities in Musicians Nadine Pecenka, *1 Peter E. Keller, *2 * Music Cognition and Action Group, Max Planck Institute for Human Cognitive
Expressive Singing Synthesis based on Unit Selection for the Singing Synthesis Challenge 2016
 Expressive Singing Synthesis based on Unit Selection for the Singing Synthesis Challenge 2016 Jordi Bonada, Martí Umbert, Merlijn Blaauw Music Technology Group, Universitat Pompeu Fabra, Spain jordi.bonada@upf.edu,
Expressive Singing Synthesis based on Unit Selection for the Singing Synthesis Challenge 2016 Jordi Bonada, Martí Umbert, Merlijn Blaauw Music Technology Group, Universitat Pompeu Fabra, Spain jordi.bonada@upf.edu,
Automatic Laughter Detection
 Automatic Laughter Detection Mary Knox Final Project (EECS 94) knoxm@eecs.berkeley.edu December 1, 006 1 Introduction Laughter is a powerful cue in communication. It communicates to listeners the emotional
Automatic Laughter Detection Mary Knox Final Project (EECS 94) knoxm@eecs.berkeley.edu December 1, 006 1 Introduction Laughter is a powerful cue in communication. It communicates to listeners the emotional
Robert Alexandru Dobre, Cristian Negrescu
 ECAI 2016 - International Conference 8th Edition Electronics, Computers and Artificial Intelligence 30 June -02 July, 2016, Ploiesti, ROMÂNIA Automatic Music Transcription Software Based on Constant Q
ECAI 2016 - International Conference 8th Edition Electronics, Computers and Artificial Intelligence 30 June -02 July, 2016, Ploiesti, ROMÂNIA Automatic Music Transcription Software Based on Constant Q
MEASURING LOUDNESS OF LONG AND SHORT TONES USING MAGNITUDE ESTIMATION
 MEASURING LOUDNESS OF LONG AND SHORT TONES USING MAGNITUDE ESTIMATION Michael Epstein 1,2, Mary Florentine 1,3, and Søren Buus 1,2 1Institute for Hearing, Speech, and Language 2Communications and Digital
MEASURING LOUDNESS OF LONG AND SHORT TONES USING MAGNITUDE ESTIMATION Michael Epstein 1,2, Mary Florentine 1,3, and Søren Buus 1,2 1Institute for Hearing, Speech, and Language 2Communications and Digital
4. ANALOG TV SIGNALS MEASUREMENT
 Goals of measurement 4. ANALOG TV SIGNALS MEASUREMENT 1) Measure the amplitudes of spectral components in the spectrum of frequency modulated signal of Δf = 50 khz and f mod = 10 khz (relatively to unmodulated
Goals of measurement 4. ANALOG TV SIGNALS MEASUREMENT 1) Measure the amplitudes of spectral components in the spectrum of frequency modulated signal of Δf = 50 khz and f mod = 10 khz (relatively to unmodulated
Experiment 7: Bit Error Rate (BER) Measurement in the Noisy Channel
 Measurement in the Noisy Channel") Experiment 7: Bit Error Rate (BER) Measurement in the Noisy Channel Modified Dr Peter Vial March 2011 from Emona TIMS experiment ACHIEVEMENTS: ability to set up a digital communications system over a noisy,
Experiment 7: Bit Error Rate (BER) Measurement in the Noisy Channel Modified Dr Peter Vial March 2011 from Emona TIMS experiment ACHIEVEMENTS: ability to set up a digital communications system over a noisy,
Brain-Computer Interface (BCI)
") Brain-Computer Interface (BCI) Christoph Guger, Günter Edlinger, g.tec Guger Technologies OEG Herbersteinstr. 60, 8020 Graz, Austria, guger@gtec.at This tutorial shows HOW-TO find and extract proper signal
Brain-Computer Interface (BCI) Christoph Guger, Günter Edlinger, g.tec Guger Technologies OEG Herbersteinstr. 60, 8020 Graz, Austria, guger@gtec.at This tutorial shows HOW-TO find and extract proper signal
PRELIMINARY INFORMATION. Professional Signal Generation and Monitoring Options for RIFEforLIFE Research Equipment
 Integrated Component Options Professional Signal Generation and Monitoring Options for RIFEforLIFE Research Equipment PRELIMINARY INFORMATION SquareGENpro is the latest and most versatile of the frequency
Integrated Component Options Professional Signal Generation and Monitoring Options for RIFEforLIFE Research Equipment PRELIMINARY INFORMATION SquareGENpro is the latest and most versatile of the frequency
SUPPLEMENTARY INFORMATION
 doi: 1.138/nature691 SUPPLEMENTAL METHODS Chronically Implanted Electrode Arrays Warp16 electrode arrays (Neuralynx Inc., Bozeman MT) were used for these recordings. These arrays consist of a 4x4 array
doi: 1.138/nature691 SUPPLEMENTAL METHODS Chronically Implanted Electrode Arrays Warp16 electrode arrays (Neuralynx Inc., Bozeman MT) were used for these recordings. These arrays consist of a 4x4 array
Practicum 3, Fall 2010
 A. F. Miller 2010 T1 Measurement 1 Practicum 3, Fall 2010 Measuring the longitudinal relaxation time: T1. Strychnine, dissolved CDCl3 The T1 is the characteristic time of relaxation of Z magnetization
A. F. Miller 2010 T1 Measurement 1 Practicum 3, Fall 2010 Measuring the longitudinal relaxation time: T1. Strychnine, dissolved CDCl3 The T1 is the characteristic time of relaxation of Z magnetization
1 Introduction to PSQM
 A Technical White Paper on Sage s PSQM Test Renshou Dai August 7, 2000 1 Introduction to PSQM 1.1 What is PSQM test? PSQM stands for Perceptual Speech Quality Measure. It is an ITU-T P.861 [1] recommended
A Technical White Paper on Sage s PSQM Test Renshou Dai August 7, 2000 1 Introduction to PSQM 1.1 What is PSQM test? PSQM stands for Perceptual Speech Quality Measure. It is an ITU-T P.861 [1] recommended
Musicians Adjustment of Performance to Room Acoustics, Part III: Understanding the Variations in Musical Expressions
 Musicians Adjustment of Performance to Room Acoustics, Part III: Understanding the Variations in Musical Expressions K. Kato a, K. Ueno b and K. Kawai c a Center for Advanced Science and Innovation, Osaka
Musicians Adjustment of Performance to Room Acoustics, Part III: Understanding the Variations in Musical Expressions K. Kato a, K. Ueno b and K. Kawai c a Center for Advanced Science and Innovation, Osaka
Limitations of a Load Pull System
 Limitations of a Load Pull System General Rule: The Critical Sections in a Load Pull measurement setup are the sections between the RF Probe of the tuners and the DUT. The Reflection and Insertion Loss
Limitations of a Load Pull System General Rule: The Critical Sections in a Load Pull measurement setup are the sections between the RF Probe of the tuners and the DUT. The Reflection and Insertion Loss
Sound Quality Analysis of Electric Parking Brake
 Sound Quality Analysis of Electric Parking Brake Bahare Naimipour a Giovanni Rinaldi b Valerie Schnabelrauch c Application Research Center, Sound Answers Inc. 6855 Commerce Boulevard, Canton, MI 48187,
Sound Quality Analysis of Electric Parking Brake Bahare Naimipour a Giovanni Rinaldi b Valerie Schnabelrauch c Application Research Center, Sound Answers Inc. 6855 Commerce Boulevard, Canton, MI 48187,
A Real Word Case Study E- Trap by Bag End Ovasen Studios, New York City
 21 March 2007 070315 - dk v5 - Ovasen Case Study Written by David Kotch Edited by John Storyk A Real Word Case Study E- Trap by Bag End Ovasen Studios, New York City 1. Overview - Description of Problem
21 March 2007 070315 - dk v5 - Ovasen Case Study Written by David Kotch Edited by John Storyk A Real Word Case Study E- Trap by Bag End Ovasen Studios, New York City 1. Overview - Description of Problem
A Real Word Case Study E- Trap by Bag End Ovasen Studios, New York City
 21 March 2007 070315 - dk v5 - Ovasen Case Study Written by David Kotch Edited by John Storyk A Real Word Case Study E- Trap by Bag End Ovasen Studios, New York City 1. Overview - Description of Problem
21 March 2007 070315 - dk v5 - Ovasen Case Study Written by David Kotch Edited by John Storyk A Real Word Case Study E- Trap by Bag End Ovasen Studios, New York City 1. Overview - Description of Problem
SYNTHESIS FROM MUSICAL INSTRUMENT CHARACTER MAPS
 Published by Institute of Electrical Engineers (IEE). 1998 IEE, Paul Masri, Nishan Canagarajah Colloquium on "Audio and Music Technology"; November 1998, London. Digest No. 98/470 SYNTHESIS FROM MUSICAL
Published by Institute of Electrical Engineers (IEE). 1998 IEE, Paul Masri, Nishan Canagarajah Colloquium on "Audio and Music Technology"; November 1998, London. Digest No. 98/470 SYNTHESIS FROM MUSICAL
DP1 DYNAMIC PROCESSOR MODULE OPERATING INSTRUCTIONS
 DP1 DYNAMIC PROCESSOR MODULE OPERATING INSTRUCTIONS and trouble-shooting guide LECTROSONICS, INC. Rio Rancho, NM INTRODUCTION The DP1 Dynamic Processor Module provides complete dynamic control of signals
DP1 DYNAMIC PROCESSOR MODULE OPERATING INSTRUCTIONS and trouble-shooting guide LECTROSONICS, INC. Rio Rancho, NM INTRODUCTION The DP1 Dynamic Processor Module provides complete dynamic control of signals
inter.noise 2000 The 29th International Congress and Exhibition on Noise Control Engineering August 2000, Nice, FRANCE
 Copyright SFA - InterNoise 2000 1 inter.noise 2000 The 29th International Congress and Exhibition on Noise Control Engineering 27-30 August 2000, Nice, FRANCE I-INCE Classification: 7.9 THE FUTURE OF SOUND
Copyright SFA - InterNoise 2000 1 inter.noise 2000 The 29th International Congress and Exhibition on Noise Control Engineering 27-30 August 2000, Nice, FRANCE I-INCE Classification: 7.9 THE FUTURE OF SOUND
The Healing Power of Music. Scientific American Mind William Forde Thompson and Gottfried Schlaug
 The Healing Power of Music Scientific American Mind William Forde Thompson and Gottfried Schlaug Music as Medicine Across cultures and throughout history, music listening and music making have played a
The Healing Power of Music Scientific American Mind William Forde Thompson and Gottfried Schlaug Music as Medicine Across cultures and throughout history, music listening and music making have played a
2 MHz Lock-In Amplifier
 2 MHz Lock-In Amplifier SR865 2 MHz dual phase lock-in amplifier SR865 2 MHz Lock-In Amplifier 1 mhz to 2 MHz frequency range Dual reference mode Low-noise current and voltage inputs Touchscreen data display
2 MHz Lock-In Amplifier SR865 2 MHz dual phase lock-in amplifier SR865 2 MHz Lock-In Amplifier 1 mhz to 2 MHz frequency range Dual reference mode Low-noise current and voltage inputs Touchscreen data display
WHAT MAKES FOR A HIT POP SONG? WHAT MAKES FOR A POP SONG?
 WHAT MAKES FOR A HIT POP SONG? WHAT MAKES FOR A POP SONG? NICHOLAS BORG AND GEORGE HOKKANEN Abstract. The possibility of a hit song prediction algorithm is both academically interesting and industry motivated.
WHAT MAKES FOR A HIT POP SONG? WHAT MAKES FOR A POP SONG? NICHOLAS BORG AND GEORGE HOKKANEN Abstract. The possibility of a hit song prediction algorithm is both academically interesting and industry motivated.
The Power of Listening
 The Power of Listening Auditory-Motor Interactions in Musical Training AMIR LAHAV, a,b ADAM BOULANGER, c GOTTFRIED SCHLAUG, b AND ELLIOT SALTZMAN a,d a The Music, Mind and Motion Lab, Sargent College of
The Power of Listening Auditory-Motor Interactions in Musical Training AMIR LAHAV, a,b ADAM BOULANGER, c GOTTFRIED SCHLAUG, b AND ELLIOT SALTZMAN a,d a The Music, Mind and Motion Lab, Sargent College of
Simple motion control implementation
 Simple motion control implementation with Omron PLC SCOPE In todays challenging economical environment and highly competitive global market, manufacturers need to get the most of their automation equipment
Simple motion control implementation with Omron PLC SCOPE In todays challenging economical environment and highly competitive global market, manufacturers need to get the most of their automation equipment
Musical Illusions Diana Deutsch Department of Psychology University of California, San Diego La Jolla, CA 92093
 Musical Illusions Diana Deutsch Department of Psychology University of California, San Diego La Jolla, CA 92093 ddeutsch@ucsd.edu In Squire, L. (Ed.) New Encyclopedia of Neuroscience, (Oxford, Elsevier,
Musical Illusions Diana Deutsch Department of Psychology University of California, San Diego La Jolla, CA 92093 ddeutsch@ucsd.edu In Squire, L. (Ed.) New Encyclopedia of Neuroscience, (Oxford, Elsevier,
Advanced Test Equipment Rentals ATEC (2832)
") E stablished 1981 Advanced Test Equipment Rentals www.atecorp.com 800-404-ATEC (2832) Technical Datasheet Scalar Network Analyzer Model 8003-10 MHz to 40 GHz The Giga-tronics Model 8003 Precision Scalar
E stablished 1981 Advanced Test Equipment Rentals www.atecorp.com 800-404-ATEC (2832) Technical Datasheet Scalar Network Analyzer Model 8003-10 MHz to 40 GHz The Giga-tronics Model 8003 Precision Scalar
LOUDNESS EFFECT OF THE DIFFERENT TONES ON THE TIMBRE SUBJECTIVE PERCEPTION EXPERIMENT OF ERHU
 The 21 st International Congress on Sound and Vibration 13-17 July, 2014, Beijing/China LOUDNESS EFFECT OF THE DIFFERENT TONES ON THE TIMBRE SUBJECTIVE PERCEPTION EXPERIMENT OF ERHU Siyu Zhu, Peifeng Ji,
The 21 st International Congress on Sound and Vibration 13-17 July, 2014, Beijing/China LOUDNESS EFFECT OF THE DIFFERENT TONES ON THE TIMBRE SUBJECTIVE PERCEPTION EXPERIMENT OF ERHU Siyu Zhu, Peifeng Ji,