Department of Psychology, University of York. NIHR Nottingham Hearing Biomedical Research Unit. Hull York Medical School, University of York
|
|
|
- Oliver Patrick
- 5 years ago
- Views:
Transcription
1 Peripheral hearing loss reduces the ability of 2 children to direct selective attention during multi- 3 talker listening 4 Emma Holmes a, Padraig T. Kitterick b, and A. Quentin Summerfield a, c 5 a Department of Psychology, University of York 6 b NIHR Nottingham Hearing Biomedical Research Unit 7 c Hull York Medical School, University of York 8 9 Corresponding author: Emma Holmes 1, eholme5@uwo.ca 1 Current postal address: The Brain and Mind Institute, Natural Sciences Centre, Room 120, Western University, London, N6A 5B7 Ontario, Canada 1
2 Abstract 2 Restoring normal hearing requires knowledge of how peripheral and central auditory processes are 3 affected by hearing loss. Previous research has focussed primarily on peripheral changes following 4 sensorineural hearing loss, whereas consequences for central auditory processing have received less 5 attention. We examined the ability of hearing-impaired children to direct auditory attention to a voice 6 of interest (based on the talker s spatial location or gender) in the presence of a common form of 7 background noise: the voices of competing talkers (i.e. during multi-talker, or Cocktail Party 8 listening). We measured brain activity using electro-encephalography (EEG) when children prepared 9 to direct attention to the spatial location or gender of an upcoming target talker who spoke in a 10 mixture of three talkers. Compared to normally-hearing children, hearing-impaired children showed 11 significantly less evidence of preparatory brain activity when required to direct spatial attention. This 12 finding is consistent with the idea that hearing-impaired children have a reduced ability to prepare 13 spatial attention for an upcoming talker. Moreover, preparatory brain activity was not restored when 14 hearing-impaired children listened with their acoustic hearing aids. An implication of these findings is 15 that steps to improve auditory attention alongside acoustic hearing aids may be required to improve 16 the ability of hearing-impaired children to understand speech in the presence of competing talkers. 17 Key words 18 Hearing loss; Multi-talker listening; Auditory Attention; Spatial attention; EEG; CNV 2
3 Introduction 2 Listeners with normal hearing can deploy attention successfully and flexibly to a talker of 3 interest when multiple talkers speak at the same time (Larson and Lee, 2014; O Sullivan et al., 2014), 4 an ability that is fundamental to successful verbal communication. These multi-talker (or Cocktail 5 Party ) listening environments are particularly challenging for people with hearing loss, as 6 demonstrated both by accuracy scores and self-report (Dubno et al., 1984; Helfer and Freyman, 2008). 7 As a result of this difficulty, children with hearing loss may be at a particular disadvantage when 8 learning language, because they not only have to do so with distorted representations of the acoustic 9 features of speech, but also frequently hear speech in acoustic environments with multiple competing 10 talkers. At least part of the difficulty in multi-talker listening arises from impairments in peripheral 11 transduction in the ear, including loss of sensitivity to higher frequencies (Hogan and Turner, 1998), 12 impaired frequency selectivity (Gaudrain et al., 2007; Moore, 1998), and impaired ability to interpret 13 temporal fine structure (Lorenzi et al., 2006). However, it is currently unclear to what extent atypical 14 cognitive abilities contribute to the difficulties in multi-talker listening experienced by children with 15 moderate hearing loss (who experience distortions in peripheral processing, although retain residual 16 hearing). The current experiments compared the ability of hearing-impaired and normally-hearing 17 children to direct preparatory attention to the spatial location or gender of a talker during multi-talker 18 listening. 19 Cognitive abilities have been found to differ between children with normal hearing and 20 children who use cochlear implants (CIs). Children with severe-to-profound loss who use CIs score 21 more poorly on tests of working memory and inhibitory control than normally-hearing children (Beer 22 et al., 2014, 2011). This finding demonstrates that atypical auditory input can potentially affect the 23 development of cognitive abilities. However, the extent to which preserved auditory encoding matters 24 for executive function is currently unclear. Given that children with CIs have minimal residual hearing 25 and may have undergone a period of auditory deprivation in childhood prior to implantation, it is 3
4 unclear whether adults who acquired hearing loss later in life or people with less severe hearing losses 2 would also exhibit atypical executive functions. 3 As a result of the inherent difficulty of separating peripheral from cognitive processes, it 4 remains unclear whether moderate hearing loss has downstream consequences for cognitive auditory 5 abilities. Neher et al. (2009) used the Test of Everyday Attention (Robertson et al., 1996) to measure 6 attention and working memory in adults with moderate hearing loss. Speech reception thresholds in 7 hearing-impaired adults during multi-talker listening were correlated with selective attention, 8 attentional switching, and working memory. However, most of the participants were older adults 9 (mean age of 60 years) and speech reception thresholds were significantly correlated with age; thus, 10 it is possible that declines in cognitive and peripheral auditory processing are unrelated to each other, 11 but both related independently to aging (for example, as a result of decreased cortical volume in older 12 people; e.g. Cardin, 2016). 13 Instead of using behavioural tests to investigate cognitive function, several studies have 14 measured cortical responses in listeners with moderate hearing loss. For example, Peelle et al. (2011) 15 found that average pure-tone hearing thresholds predicted the extent to which spoken sentences 16 evoked activity in the bilateral superior temporal gyri, thalamus, and brainstem in hearing-impaired 17 adults. Several studies using electro-encephalography (EEG) and magneto-encephalography (MEG) 18 have also shown atypical auditory evoked activity in hearing-impaired adults (Alain et al., 2014; 19 Campbell and Sharma, 2013; Oates et al., 2002) and children (Koravand et al., 2012). However, 20 although these studies measured cortical activity, they do not necessarily indicate atypical cognitive 21 processes in hearing-impaired listeners: differences in neural activity between normally-hearing and 22 hearing-impaired listeners could arise either due to impaired cognitive function or because normal 23 cognitive processes are deployed onto a distorted central representation of the acoustic signal. The 24 current experiment avoided this confound by seeking evidence of differences in neural activity when 25 participants prepared to direct attention to speech (i.e. before the speech began) during multi-talker 26 listening. 4
5 Normally-hearing listeners can use between-talker differences in acoustic properties as cues 2 to improve the intelligibility of speech spoken by a target talker during multi-talker listening. For 3 example, normally-hearing listeners show better speech intelligibility when the talkers differ in gender 4 (Brungart, 2001; Brungart et al., 2001; Shafiro and Gygi, 2007), fundamental frequency (Assmann and 5 Summerfield, 1994; Darwin and Hukin, 2000), or spatial location (Bronkhorst and Plomp, 1988; Darwin 6 and Hukin, 1999; Helfer and Freyman, 2005). Normally-hearing listeners can also deploy preparatory 7 attention to these acoustic cues before a target talker starts to speak. First, they achieve better 8 accuracy of speech intelligibility when they know the spatial location (Best et al., 2009, 2007; Ericson 9 et al., 2004; Kidd et al., 2005) or the identity (Freyman et al., 2004; Kitterick et al., 2010) of a target 10 talker before he or she begins to speak. Second, previous experiments using functional magnetic 11 resonance imaging (fmri; Hill and Miller, 2010) and MEG (Lee et al., 2013) have revealed preparatory 12 brain activity that differs depending on whether normally-hearing adults direct attention to the spatial 13 location or fundamental frequency of the target talker. Normally-hearing adults and children also 14 show preparatory EEG activity when they are cued to the location or gender of a target talker (Holmes 15 et al., 2016). If hearing-impaired children deploy preparatory attention in a similar way as normally- 16 hearing children do, there should be no differences in preparatory EEG activity between normally- 17 hearing and hearing-impaired children. 18 In the current experiment, we presented an adult male and an adult female voice concurrently 19 from different spatial locations. A third, child s, voice was also presented to increase the difficulty of 20 the task. Prior to the presentation of the voices, a visual stimulus cued attention to either the spatial 21 location or gender of the target talker, who was always one of the two adults. The task was to report 22 key words spoken by the target talker. We recorded brain activity using electro-encephalography 23 (EEG) in children with moderate sensorineural hearing loss of several year s duration (HI children) and 24 in a comparison group of normally-hearing (NH) children. We isolated preparatory EEG activity by 25 comparing event-related potentials (ERPs) between a condition in which the visual cue indicated the 26 location or gender of an upcoming target talker and a control condition in which the same visual cues 5
6 were presented but did not instruct participants to attend to acoustic stimuli. We hypothesised that 2 we would find less evidence of preparatory EEG activity in hearing-impaired children than in normally- 3 hearing children Methods Participants 6 Participants were 24 children with normal hearing (9 male), aged 8 15 years (mean [M] = 12.3, 7 standard deviation [SD] = 1.9) and 14 children with sensorineural hearing loss (4 male), aged years (M = 11.6, SD = 3.1). All participants were declared by their parents to be native English speakers. 9 The NH children were all also declared by their parents to be right-handed with no history of hearing 10 problems and they had 5-frequency average pure-tone hearing levels of 15 db HL or better, tested in 11 accordance with BS EN ISO (British Society of Audiology, 2004; Fig. 1). The children with hearing 12 loss had bilateral 5-frequency average pure-tone hearing levels between 42 and 65 db HL (M = db HL, SD = 7.9; Fig. 1) and the difference in the 5-frequency averages recorded from the left and right 14 ears was less than 12 db for each participant. Of the fourteen HI children, two were left-handed and 15 one had an additional visual impairment in her left eye. The study was approved by the Research Ethics 16 Committee of the Department of Psychology, University of York, the NHS Research Ethics Committee 17 of Newcastle and North Tyneside, and the Research and Development Departments of York Teaching 18 Hospital NHS Foundation Trust, Leeds Teaching Hospitals NHS Trust, Hull and East Yorkshire Hospitals 19 NHS Trust, and Bradford Teaching Hospitals NHS Foundation Trust < Insert Fig. 1 > The HI children completed the experiment for the first time without using their hearing aids. 24 A subset of ten HI children (aged 7 16 years, M = 11.9 years, SD = 2.5; 2 male; 1 left-handed) also took 25 part in the experiment for a second time using their own acoustic bilateral behind-the-ear hearing 6
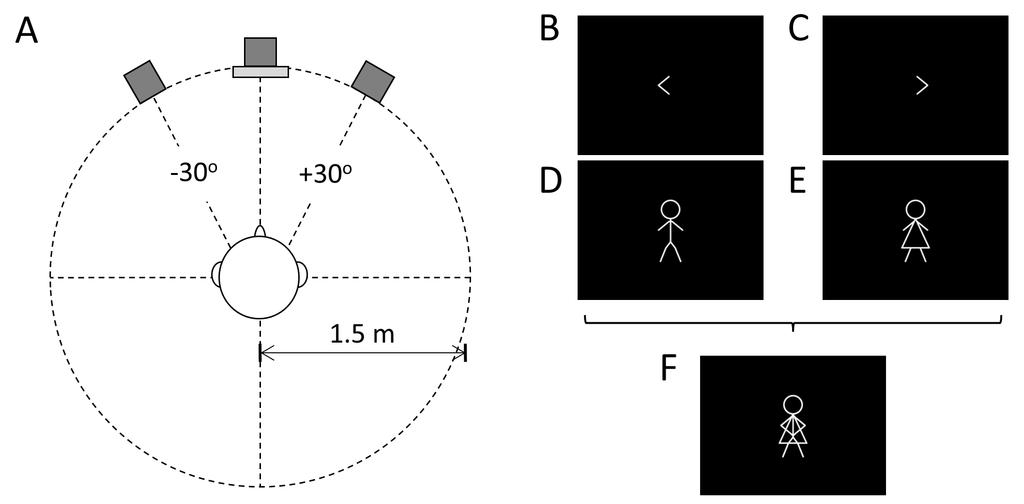
7 aids. The aided session took place between 2 and 9 months after the unaided session. We refer to the 2 entire group who participated in the unaided session as the HI U group. For the children who took part 3 in both aided and unaided sessions, we distinguish between HI A and HI U sessions, respectively Materials 5 The experiment was conducted in a 5.3 m x 3.7 m single-walled test room (Industrial Acoustics 6 Co., NY) located within a larger sound-treated room. Participants sat facing three loudspeakers (Plus 7 XS.2, Canton) arranged in a circular arc at a height of 1 m at 0 azimuth (fixation) and at 30 to the left 8 and right (Fig. 2A). A 15-inch visual display unit (VDU; NEC AccuSync 52VM) was positioned directly 9 below the central loudspeaker. 10 Four visual cues, left, right, male, and female, were defined by white lines on a black 11 background. Left and right cues were leftward- and rightward-pointing arrows, respectively; male and 12 female cues were stick figures (Fig. 2B E). A composite visual stimulus consisted of the four cues 13 overlaid (Fig. 2F) < Insert Fig. 2 > Acoustical test stimuli were modified phrases from the Co-ordinate Response Measure corpus 18 (CRM; Moore, 1981) spoken by native British English talkers (Kitterick et al., 2010). One male and one 19 female talker were selected from the corpus. An additional female talker was selected from the 20 corpus, whose voice was manipulated to sound like a child s voice by simulating a change in F0 and 21 vocal tract length using Praat (Version ; The original stimuli were edited 22 so that each phrase had the form <colour> <number> now. There were four colours ( Blue, Red, 23 Green, White ) and four numbers ( One, Two, Three, Four ). An example is Green Two now. 24 The average duration of the presented phrases was 1.4 s. The levels of the digital recordings of the 25 phrases were normalised to the same root mean square (RMS) power. 7
8 Control stimuli were single-channel noise-vocoded representations of concurrent triplets of 2 modified CRM phrases that were used as acoustical test stimuli. Each control stimulus was created by 3 summing three acoustical test phrases (one spoken by each talker) digitally with their onsets aligned 4 and extracting the temporal envelope of the combination using the Hilbert Transform (Hilbert, 1912). 5 We used the envelope to modulate the amplitude of a random noise whose long-term spectrum 6 matched the average spectrum of all of the possible triplets of phrases Procedures 8 Fig. 3A illustrates the trial structure in the test condition. The visual cue directed attention to 9 the target talker and varied quasi-randomly from trial to trial. The cue remained on the screen 10 throughout the duration of the acoustic stimuli so that participants did not have to retain the visual 11 cue in memory. The three different talkers were presented from the three loudspeakers (left, middle, 12 and right). The phrases started simultaneously, but contained different colour-number combinations. 13 The child talker was always presented from the middle loudspeaker and was always unattended. 14 Over the course of the experiment, the male and female talkers were presented equally often from 15 the left and right locations. After the phrases had ended, participants were instructed to report the 16 colour-number combination in the target phrase by pressing a coloured digit on a touch screen directly 17 in front of their chair. Each participant completed between 96 and 144 trials in the test condition 18 (depending on their stamina), with an equal number of each the four cue types. There was a short 19 break every 16 trials and longer break every 48 trials < Insert Fig. 3 > The average presentation level of concurrent pairs of test phrases was set to 63 db(a) SPL 24 (range db) for normally-hearing children and 76 db(a) SPL (range db) for 25 hearing-impaired children. This difference aimed to compensate, in part, for higher pure-tone 26 thresholds of the hearing-impaired children. Presentation levels were measured with a B&K (Brüel & 8
9 Kjær, Nærum, Denmark) Sound Level Meter (Type 2260 Investigator) and 0.5-inch Free-field 2 Microphone (Type 4189) placed in the centre of the arc at the height of the loudspeakers with the 3 participant absent. 4 The trial structure in the control condition was the same as in the test condition (Fig. 3B) with 5 the exception that an acoustical control stimulus, presented from the loudspeaker at 0 azimuth, 6 replaced the triplet of acoustical test stimuli. The purpose of the control condition was to measure 7 responses to the visual cues when they had no implications for auditory attention. The task was to 8 identify the picture that corresponded to the visual cue on each trial. The logic behind the design of 9 the control condition was that the acoustic stimuli lacked the spectral detail and temporal fine 10 structure required for the perception of pitch (Moore, 2008). In addition, because the stimuli were 11 presented from one loudspeaker, they did not provide the interaural differences in level and timing 12 required for their constituent voices to be localised separately. In these ways, the acoustic cues 13 required to segregate the sentences by gender and by location were neutralised, while the overall 14 energy and gross fluctuations in amplitude of the test stimuli were preserved. Each participant 15 completed 96 trials (24 in each cue type condition) with a short break every 12 trials and a longer 16 break every 36 trials. The presentation level of the acoustical control stimuli was set so that their 17 average level matched the average level of the triplets of test stimuli. Participants undertook the 18 control condition before the test condition; that is, before they had learnt the association between 19 the visual cues and the acoustical test stimuli. 20 After participants had completed the control condition, but before they undertook the test 21 condition, they completed two sets of familiarisation trials, which had a similar trial structure to the 22 test condition. In the first set (12 trials), either the male or female talker was presented on each trial 23 from the left or right loudspeaker. In the second set (4 trials), each trial contained all three voices, 24 identical to the test condition. EEG activity was not recorded during familiarisation Behavioural analyses 9
10 Trials were separated into location (average left/right cues) and gender (average male/female 2 cues) groups, separately for the test and control conditions. Responses were scored as correct if both 3 the colour and number key words were reported correctly in the test condition and if the visual cue 4 was reported correctly in the control condition. A 2 x 2 between-subjects ANOVA compared accuracy 5 between NH and HI U children for the location and gender cue types. A 2 x 2 within-subjects ANOVA 6 contrasted the subset of HI children who completed both the aided and unaided sessions (HI A and HI U ) EEG recording and processing 8 Continuous EEG was recorded using the ANT WaveGuard-64 system (ANT, Netherlands; 9 with Ag/AgCl electrodes (with active shielding) mounted on an elasticated cap 10 (positions: Fp1, Fp2, AF3, AF4, AF7, AF8, F1, F2, F3, F4, F5, F6, F7, F8, FC1, FC2, FC3, FC4, FC5, FC6, FT7, 11 FT8, C1, C2, C3, C4, C5, C6, T7, T8, CP1, CP2, CP3, CP4, CP5, CP6, TP7, TP8, P1, P2, P3, P4, P5, P6, P7, 12 P8, PO3, PO4, PO7, PO8, O1, O2, M1, M2, Fpz, Fz, FCz, Cz, CPz, Pz, POz, Oz). An additional electrode 13 (AFz) was used as a ground site. The horizontal electro-oculogram (EOG) was measured with a bipolar 14 lead attached to the outer canthi of the left and right eyes and the vertical EOG was measured with a 15 bipolar lead above and below the right eye. The EEG was amplified and digitised with an ANT High- 16 Speed Amplifier at a sampling rate of 1000 Hz per channel. Electrode impedances at the start of the 17 experiment were below 30 kohm. 18 The continuous EEG recordings were exported to MATLAB 7 (The MathWorks, Inc., Natick, 19 MA, USA). The data was processed using the EEGLAB toolbox (Version 9; 20 and ERPs were statistically analysed using the FieldTrip toolbox 21 ( Before statistical analysis, the data were band-pass filtered between and 30 Hz. The purpose of bandpass filtering was to remove DC offset, slow drifts due to skin 23 potentials, line noise, and muscle-related artefacts. The amplitude at each electrode was referenced 24 to the average amplitude of the electrode array. Epochs were created with 4700 ms duration, 25 including a baseline interval of 200 ms at the end of the fixation-cross period. Given that HI children 26 performed the task with low accuracy, we included correct and incorrect trials in the analyses to 10
11 improve power for detecting differences between NH and HI children. However, including incorrect 2 trials in the analysis did not lead to qualitatively different ERPs, or different conclusions from statistical 3 tests, than when incorrect trials were excluded (see Supplementary Fig. 2). Independent component 4 analysis (ICA) was used to correct for eye-blink artifacts, which were identified by a stereotyped scalp 5 topography. There were no discernible artefacts attributable to the hearing aids in the pre-processed 6 data from the HI A session Analyses of ERPs 8 Fig. 4 shows a schematic of the EEG analysis pipeline. We used cluster-based permutation 9 analyses (Maris and Oostenveld, 2007) to identify differences in EEG activity between the test and 10 control conditions (separately for location and gender trials) and between location and gender trials 11 (within the test condition). The method searches for clusters of adjacent electrodes over successive 12 time points that display systematic differences between two experimental conditions. The value of 13 the t-statistic is calculated for each electrode at each time point. Clusters are then tested for 14 significance by comparing the sum of the t-values within the observed cluster against the null 15 distribution, which is constructed by permuting the data between conditions and searching for 16 clusters in the permuted data. We used this method first to identify preparatory attention in NH 17 children and, second, in HI U children; we conducted the cluster-based permutation analysis in the 18 interval between the full reveal of the visual cue and the onset of acoustic stimuli (duration = ms) < Insert Fig. 4 > For each significant cluster identified in the NH children, the magnitude of the cluster 24 calculated as the difference in amplitude between conditions, averaged across the electrodes and 25 time points that contributed to the cluster was compared between NH and HI u children using 26 bootstrapping. First, a sample of 14 children was selected (with replacement) from the NH group; 11
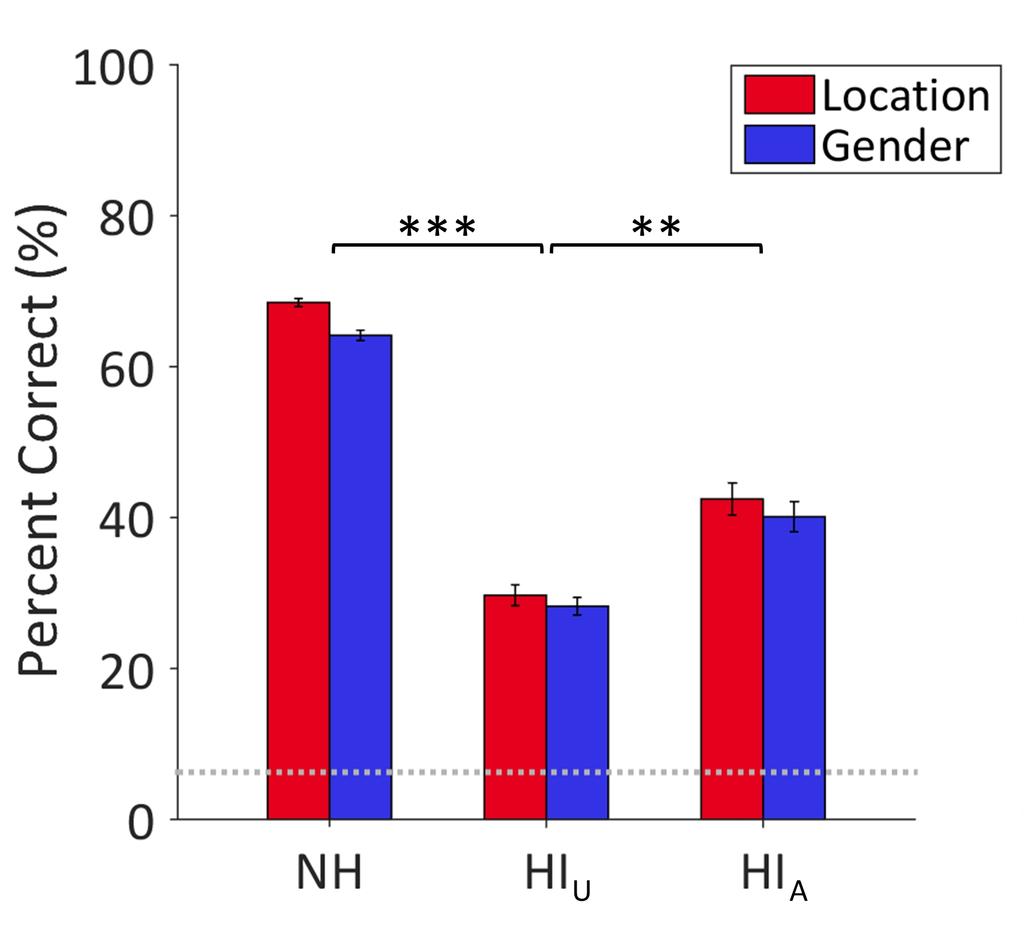
12 ,000 samples were selected to form a null distribution. Second, the average magnitude of each 2 cluster for the 14 HI U children was compared against the null distribution in a two-tailed test (ɑ = 0.95). 3 The purpose of this analysis was to equate the group sizes for NH and HI U children. The same 4 comparison was conducted between the 10 HI A children and samples of 10 NH children. 5 To compare ERPs for the hearing-impaired children when they listened aided and unaided, a 6 within-subjects t-test compared the average magnitude of each cluster in the sub-set of children who 7 completed both the aided and unaided sessions Results Behavioural results 10 NH children achieved significantly higher accuracy of speech intelligibility (M = 66.3%, SD = ) than HI U children [M = 29.0%, SD = 15.4; F(1, 36) = 51.71, p < 0.001, η 2 p = 0.59; Fig. 5], with no 12 significant difference between trials in which they were cued to location (left/right) and gender 13 (male/female) [F(1, 36) = 3.82, p = 0.06] and no significant interaction between hearing group and cue 14 type [F(1, 36) = 0.95, p = 0.34]. In the control condition, there was no significant difference in accuracy 15 for identifying the visual cues between NH (M = 98.1%, SD = 3.9) and HI u children [M = 94.7%, SD = ; F(1, 36) = 1.43, p = 0.24]. There was also no significant difference between cue types [F(1, 36) = , p = 0.09] and no significant interaction [F(1, 36) = 1.43, p = 0.24]. 18 HI children identified words spoken by the target talker with significantly higher accuracy in 19 the aided (M = 41.3%, SD = 20.4) than the unaided (M = 28.5%, SD = 20.3) session [F(1, 9) = 25.71, p = , η 2 p = 0.74]. There was no significant difference between cue types [F(1, 9) = 0.60, p = 0.46] and 21 no significant interaction [F(1, 9) = 0.92, p = 0.36]. In the control condition, there was no significant 22 difference in accuracy for identifying the visual cues between the aided (M = 93.4%, SD = 10.4) and 23 unaided (M = 94.4%, SD = 9.2) sessions [F(1, 9) = 0.38, p = 0.27] and no significant difference between 12
13 cue types [F(1, 9) = 0.16, p = 0.70]. There was a marginal significant interaction between aiding and 2 cue type in the control condition [F(1, 9) = 5.44, p = 0.045, η p 2 = 0.38]. 3 4 < Insert Fig. 5 > Event-related potentials: Evidence for preparatory attention 7 First, using cluster-based permutation analyses, we sought evidence of preparatory attention 8 in NH children. Fig. 6 illustrates the topography and time windows of clusters that showed significant 9 differences between the test and control conditions. Additional information about each cluster is 10 tabulated in Table 1. Analyses were conducted separately for trials in which participants were cued to 11 location (left/right) and gender (male/female) < Insert Fig. 6 > < Insert Table 1 > Three significant clusters of activity were found for location trials (Clusters 1 2) and one 18 significant cluster was found for gender trials (Cluster 3N). The emergence of these significant clusters 19 is compatible with the idea that NH children prepare attention for the location and gender of an 20 upcoming talker Event-related potentials: Comparisons between location and gender trials 22 To establish whether NH children showed differences in brain activity depending on the 23 attribute of the target talker to which they were attending, we compared ERPs between location and This interaction reflected average accuracy on location trials that was slightly, but not significantly, higher than on gender trials in the aided session (p = 0.40), but average accuracy that was slightly, but not significantly, higher on gender than on location trials in the unaided session (p = 0.87). 13
14 gender trials within the test condition. No significant clusters were found. Thus, further analyses 2 focussed on examining the clusters that showed significantly different activity between the test and 3 control conditions Event-related potentials: Differences between NH and HI children 5 Bootstrapping analyses compared the magnitude of each cluster between NH children and HI 6 children. Cluster magnitude was defined as the difference in amplitude between conditions, averaged 7 across the electrodes and time points that contributed to the cluster. 8 Fig. 7 illustrates the average cluster magnitude for NH and HI U children. For location trials, the 9 magnitude of all three clusters were significantly different for HI U than NH children (i.e. HI U children 10 either showed a significantly smaller difference in amplitude between the test and control conditions 11 than NH children or a difference in the opposite direction to NH children) [Cluster 1N: p = 0.002; 12 Cluster 2N: p < 0.001; Cluster 2P: p < 0.001; Table 1] < Insert Fig. 7 > Comparisons between HI A and NH children for location trials showed the same pattern of 17 results, except that the earliest cluster did not differ significantly between HI A and NH children [Cluster 18 1N: p = 0.14; Cluster 2N: p = 0.001; Cluster 2P: p = 0.002; Table 1]. 19 For gender trials, cluster magnitude did not differ significantly between NH and HI u children 20 (Cluster 3N: p = 0.13), although it did differ between NH and HI A children (Cluster 3N: p = 0.009). 21 Overall, converging results from the aided and unaided sessions show a difference in 22 preparatory EEG activity between HI and NH children during location trials (Clusters 2N and 2P) but 23 no consistent evidence for a difference during gender trials. This result demonstrates the key finding 24 that HI children prepare spatial attention to a lesser extent than NH children. 25 Additional information about each cluster is tabulated in Table 1. The ERP waveforms at each 26 cluster are illustrated in Supplementary Fig
15 Event-related potentials: Comparisons between aided and unaided conditions 2 In order to test whether aiding affected the extent of preparatory attention in HI children, the 3 magnitude of the clusters was compared between the HI U and HI A sessions. A paired-samples t-test 4 was conducted on the data from the 10 participants who completed both sessions. None of the 5 clusters showed significant differences between the aided and unaided sessions [Cluster 1N: t(9) = , p = 0.92; Cluster 2N: t(9) = 1.23, p = 0.25; Cluster 2P: t(9) = 2.13, p = 0.06; Cluster 3N: t(9) = 1.21, 7 p = 0.26]. These results suggest that different significance patterns for the comparisons of Cluster 1N 8 and 3N between NH and HI U groups and between NH and HI A groups (Section 3.4) do not reflect 9 significant differences between aided and unaided listening. The results demonstrate that aiding did 10 not affect magnitude of the clusters; thus, there was no greater evidence of preparatory attention in 11 HI children when they used their hearing aids than when they listened unaided Event-related potentials: Clusters in HI children 13 To investigate whether HI and NH children showed qualitatively different patterns of brain 14 activity, we also conducted spatio-temporal cluster-based permutation analyses on the data from the 15 HI U children, without limiting the analyses to specific groups of electrodes or time points. In other 16 words, these further analyses aimed to determine whether the group of HI children showed consistent 17 evidence of preparatory attention (indicated by the presence of a significant spatio-temporal cluster) 18 that differed in magnitude from activity in NH children. 19 We found no significant clusters for location trials (Fig. 8A). One significant cluster was found 20 for gender trials, which occurred soon after the visual cue was revealed (Cluster 4N; Fig. 8B C; Table 21 1). We compared the magnitude of this cluster between NH and HI U children in a bootstrapping 22 analysis, using the method described in Section 3.4. There was no significant difference in the 23 magnitude of Cluster 4N between NH (M = μv) and HI U (M = μv) children (p = 0.08; Fig. 24 8D), suggesting that HI children did not evoke qualitatively different EEG activity to NH children < Insert Fig. 8 > 15
16 Event-related potentials: Variability in NH and HI children 3 Given our sample of HI children varied in both age and aetiology, it was possible that the HI 4 children were more variable in evoking preparatory EEG activity than NH children. We used Levene s 5 test for equality of variances to determine whether the variance in cluster magnitude differed 6 between the NH and HI U children. There were no significant differences in variance for any of the four 7 clusters found in NH children [Cluster 1N: F = 0.70, p = 0.41; Cluster 2N: F = 27, p = 0.61; Cluster 2P: F 8 = 0.26, p = 0.61; Cluster 3N: F = 2.67, p = 0.11]. This result demonstrates that HI children were no more 9 variable than NH children in evoking preparatory EEG activity. Thus, increased variability was not the 10 reason why we found fewer significant clusters in HI children than NH children Discussion 12 HI children showed significantly less evidence of preparatory attention than NH children, 13 demonstrated by smaller differences in event-related potentials (ERPs) when visual stimuli cued 14 spatial attention to one of three talkers compared to when the same visual stimuli had no implications 15 for auditory attention. Such differences would arise if hearing-impaired children deployed less 16 preparatory activity than normally-hearing children, or if they invoked activity with different latencies 17 or in different brain regions that varied across the group of hearing-impaired children. Thus, the result 18 is compatible with the idea that HI children prepare spatial attention less consistently than NH 19 children Preparatory EEG activity in NH children 21 Previous experiments demonstrate that adults and children aged 7 13 years with normal 22 hearing show preparatory brain activity before a target talker begins to speak (Hill and Miller, 2010; 23 Holmes et al., 2016; Lee et al., 2013). Consistent with this finding, NH children aged 8 15 years in the 24 current experiment showed significant differences in ERPs between trials in which a visual cue directed 25 attention to the spatial location of an upcoming talker and trials in which the same visual cue was 16
17 presented but did not have implications for auditory attention. The current results are consistent with 2 the idea that NH children prepare their attention for the location of an upcoming target talker during 3 multi-talker listening. 4 Preparation for location evoked significant activity in two distinct time periods: the first 5 started shortly (< 75 ms) after the visual cue was revealed and lasted for approximately 300 ms; the 6 second occurred throughout the 1000 ms immediately before the talkers began to speak. In general, 7 these findings are consistent with the idea that participants with normal hearing evoke preparatory 8 brain activity before the onset of an acoustical target stimulus (Banerjee et al., 2011; Müller and Weisz, ; Voisin et al., 2006). These findings are also consistent with the results of previous experiments 10 with a similar design that tested adults and children with normal hearing (Holmes et al., 2016). Holmes 11 et al. (2016) used a speech intelligibility task that was similar to the current experiment, except that 12 (1) two, rather than three, talkers spoke simultaneously and (2) the preparatory interval was 1000 ms 13 instead of 2000 ms. Similar to the current experiment, Holmes et al. (2016) found preparatory activity 14 that began soon after a visual cue for location was presented and which was sustained before two 15 talkers started speaking. However, by using a longer preparatory interval, the current experiment 16 separated preparatory activity that occurred in two distinct time periods: the first occurred shortly 17 after the visual cue was revealed and thus likely reflects initial processing and interpretation of the 18 cue; the second occurred immediately before the talkers begin speaking and may therefore reflect 19 anticipation of characteristics of the upcoming talkers. 20 The preparatory ERPs identified in NH children that occurred in the 1000 ms immediately 21 before the talkers began to speak resemble the contingent negative variation (CNV; Walter et al., ), an ERP thought to reflect anticipation of an upcoming stimulus (e.g. Chennu et al., 2013). 23 Figures 6C (location trials) and 6F (gender trials) show that ERPs in the test condition were significantly 24 more negative than the control condition immediately before the talkers started speaking ( ms 25 prior to the onset of the talkers in location trials and ms prior in gender trials); during these 26 time periods, ERPs elicited by visual cues in the control condition (in which acoustic stimuli were 17
18 presented but were not relevant to the participants task) were approximately at baseline level, 2 whereas ERPs in the test condition were negative. Thus, differences in ERPs between the test and 3 control conditions in Figures 6C and 6F might possibly reflect the CNV (although it is unclear whether 4 the topography observed in the current experiment matches that of the CNV, given that the current 5 experiment used the average reference and previous CNV experiments have typically used a mastoids 6 or tip of the nose reference). 7 The latency of the CNV is correlated with the length of subjective judgements of interval 8 duration (Ruchkin et al., 1977), suggesting that the CNV reflects anticipation of the time at which a 9 target stimulus will occur. In addition, the CNV has been observed in both the visual and auditory 10 modalities (e.g. Pasinski et al., 2016; Walter et al., 1964), which suggests it reflects preparation that is 11 not specific to any particular attribute or modality. Indeed, consistent with the idea that the CNV does 12 not only reflect preparation for one particular stimulus attribute, we observed activity resembling the 13 CNV on both location (Figure 6C) and gender (Figure 6F) trials and found no significant differences in 14 preparatory ERPs between location and gender trials. Given that larger CNV magnitudes are related 15 to better detection of acoustic target stimuli (Rockstroh et al., 1993), the activity shown in Figures 6C 16 and 6F may reflect preparatory activity that is beneficial for speech intelligibility during multi-talker 17 listening Differences between NH and HI children 19 Comparisons between NH and HI children showed atypical ERPs in HI children during location 20 trials the difference in amplitude between the test and control conditions was significantly smaller 21 for HI than NH children (Clusters 2N and 2P; Fig. 7A). Moreover, that result was found when HI children 22 listened both unaided and aided. This result is consistent with the idea that HI children do not deploy 23 preparatory spatial attention to the same extent as NH children. Compatible with this finding, HI 24 children also showed significantly poorer accuracy of speech intelligibility than NH children. Since 25 directing preparatory spatial attention has previously been found to improve the understanding of a 26 talker by adults with normal hearing (Best et al., 2007; Ericson et al., 2004; Kidd et al., 2005), it is 18
19 possible that difficulties preparing spatial attention contributed to poor speech understanding by HI 2 children during the current task. The idea that HI children do not engage preparatory brain activity to 3 the same extent as NH children is consistent with the results of Best et al. (2009) who showed that 4 adults with moderate hearing loss gained less improvement in the accuracy of speech intelligibility 5 than NH adults when they were cued to the spatial location of a talker. Together, the findings of Best 6 et al. and the current experiment suggest that hearing loss leads to atypical preparatory attention, 7 which reduces the benefit to speech understanding gained from knowing the spatial location of a 8 talker before they start speaking. 9 One difference between HI and NH children was in the cluster that resembled the CNV (Cluster 10 2N, Figure 7A). There is some evidence from magnetoencephalography (MEG; Basile et al., 1997) and 11 EEG (Segalowitz and Davies, 2004) source localisation that the magnitude of the CNV is related to the 12 magnitude of activity in prefrontal cortex. Segalowitz and Davis (2004) showed that the development 13 of executive functions, such as working memory, in children relates to the strength of the CNV in a 14 Go/No-Go task and they, thus, suggest that the CNV may relate to development of the frontal 15 attentional network. Consistent with this idea, lower CNV magnitudes are observed in reaction-time 16 tasks when distracting visual stimuli that need to later be recalled are presented in the interval 17 between a cue and an auditory target stimulus than when no distracting stimuli are presented (Tecce 18 and Scheff, 1969; Travis and Tecce, 1998). Thus, it is possible that the difference in Cluster 2N between 19 HI and NH children could result from HI children having a less mature frontal attentional network. On 20 the other hand, Wӧstmann et al. (2015) showed that, within participants, the magnitude of the CNV 21 related to task difficulty and to the extent of temporal fine structure degradation of acoustic speech 22 stimuli. Therefore, the difference in Cluster 2N in the current experiment could reflect greater 23 difficulty of multi-talker listening for HI children, a loss of temporal fine structure information resulting 24 from hearing loss, or a combination of both of these factors. Future experiments could distinguish 25 these possibilities by examining the extent to which the difference in preparatory ERPs exists between 26 NH and HI children under different task conditions. For example, preparatory brain activity could be 19
20 compared between NH and HI children during multi-talker listening when the speech stimuli are 2 degraded for both groups and when the accuracy of speech intelligibility is similar for NH and HI 3 children. Any differences in preparatory brain activity could attempt to be localised using EEG or MEG 4 source reconstruction techniques to examine whether differences could be attributable to 5 development of the frontal attention network. 6 The current results demonstrate atypical spatial auditory attention in children with moderate 7 hearing loss, although the typical role of experience on the development of this ability is unclear. One 8 hypothesis is that a degraded representation of the cues used to distinguish talkers by their location 9 results in a reduced ability to prepare to attend to a talker based on his or her spatial location. This 10 hypothesis is consistent with the idea that reduced preparatory spatial attention is a direct 11 consequence of hearing loss and predicts that atypical spatial attention would be observed in all 12 listeners whose hearing loss distorts the ability to resolve sounds at different spatial locations. In 13 addition, this hypothesis suggests that preparatory spatial attention could be restored only if the 14 peripheral representation of spatial location is also restored. Alternatively, hearing loss may affect the 15 ability to direct selective attention in a more general manner that is not specific to the peripheral cues 16 to which the listener has access. The latter hypothesis seems more likely, given that hearing-impaired 17 children in the current experiment were able to perform the task with above-chance accuracy despite 18 showing no consistent evidence of preparatory attention. This result suggests that the children had 19 sufficient peripheral representations of spatial location to identify a target talker based on their 20 location. However, further work is required to disambiguate these two alternatives. For example, 21 future experiments could investigate the relationship between spatial localisation and/or 22 discrimination abilities and preparatory attention in hearing-impaired people. 23 During gender trials, there was no consistent evidence for atypical ERPs in HI children, 24 although, NH children did not display preparatory attention for gender to the same extent as they 25 displayed preparatory attention for location (Fig. 7). It is possible that the cues for gender used in the 26 current experiment evoked preparatory attention only minimally for both NH and HI children. This 20
21 interpretation is consistent with the results of Holmes et al. (2016) who also found minimal evidence 2 of preparatory EEG activity when NH children were cued to the gender of a target talker. 3 The analyses reported in this paper included correct and incorrect trials. The rationale was 4 that HI children performed the task with low accuracy and, therefore, removing all incorrect trials 5 would lead to lower signal-to-noise ratio in the average ERPs and, hence, lower statistical power to 6 detect differences between NH and HI children. However, this decision meant that differences in EEG 7 activity between NH and HI children could potentially reflect differences in behavioural performance 8 between NH and HI children, rather than the EEG activity that accompanied successful trials (which 9 might produce confounds, for example, if one group was not engaged in the task for all trials of the 10 experiment). We, thus, conducted a separate analysis in HI children comparing activity evoked on 11 correct trials with average activity evoked on correct and incorrect trials. The analysis of correct trials 12 revealed similar patterns of activity as the analysis that included correct and incorrect trials. This result 13 suggests that differences between NH and HI children cannot be explained by the contribution of 14 qualitatively different activity on incorrect than correct trials. Instead, the results are attributable to 15 differences in preparatory EEG activity between the NH and HI groups Effect of aiding 17 A within-subjects comparison between the aided and unaided sessions (which were 18 conducted on different days, separated by up to nine months) showed no significant difference in the 19 magnitude of the clusters. In addition, comparisons between NH and HI A groups showed similar results 20 to comparisons between NH and HI U children in both instances, Clusters 2N and 2P (which occurred 21 on location trials) showed significant differences between the NH and HI children. This result 22 demonstrates that differences in preparatory attention between HI and NH children did not arise due 23 to unfamiliar listening conditions or lack of audibility in the HI children. Another implication of this 24 result is that acoustic hearing aids do not restore normal preparatory spatial attention in children with 25 moderate sensorineural hearing loss Possible compensatory mechanisms 21
22 The results demonstrate that HI children do not display the same preparatory processes as 2 NH children when they are cued to the location of an upcoming talker. Furthermore, we found no 3 consistent evidence of preparatory spatial attention in HI children because there were no significant 4 clusters in HI children during location trials (Fig. 8A). This outcome is consistent with the idea that HI 5 children did not systematically compensate for hearing loss by engaging qualitatively different 6 preparatory brain activity to NH children or by engaging similar brain activity with a different time 7 course. Rather, the results are consistent with the idea that the group of HI children, overall, showed 8 either weaker or less consistent preparatory spatial attention than the group of NH children. 9 There was one significant cluster in HI children during gender trials, which occurred very soon 10 after the visual cue was revealed (Fig. 8B C). However, there was no evidence that the magnitude of 11 this cluster differed between NH and HI U children, which is again consistent with idea that HI children 12 did not engage qualitatively different preparatory brain activity to NH children. 13 Although HI children did not show additional preparatory activity that was different to the NH 14 children, different hearing-impaired children might have adopted different strategies to prepare 15 attention. The resulting lack of consistency might explain the general absence of significant clusters in 16 the group of HI children. We do not have information about the specific aetiology, duration of hearing 17 loss, or time of onset of the hearing loss for the HI children, but variability in these factors could 18 potentially be related to differences in preparatory attention. On the other hand, if those factors had 19 a large impact on preparatory EEG activity, we would expect individual variability in HI children to be 20 greater than that in NH children. The data do not provide evidence to support this idea, given that the 21 variance in cluster magnitude did not differ significantly between HI U and NH children. Although the 22 current numbers of participants do not provide sufficient power to examine whether preparatory EEG 23 activity related to age or audiometric thresholds, characterising the factors that influence the extent 24 of preparatory attention in children with normal and impaired hearing would be an interesting aim for 25 future studies. 22
23 The children who took part in the current experiment may have undergone a period of 2 auditory deprivation resulting from their hearing loss during a critical or sensitive period of 3 development. If this explanation is correct, individuals who acquired hearing loss during adulthood 4 may not show similar deficits in preparatory attention. Furthermore, preparatory attention would be 5 expected to differ between different people with hearing loss, depending on the age of onset of their 6 hearing loss and perhaps also on the age at which they received hearing aids. 7 In addition, the current experiment tested individuals with moderate hearing loss and, thus, 8 it is not clear whether the extent of hearing loss affects the extent to which attention is atypical. Beer 9 et al. (2011, 2014) measured executive functions in children with severe-to-profound hearing loss who 10 used CIs. Compared to normally-hearing children, children with CIs showed reduced ability to perform 11 tests of working memory and inhibitory control. This result is consistent with the idea that hearing 12 loss has consequences for central processing. This result is also relevant to the current findings 13 because preparing to attend to a talker may be related to the processes of maintaining in memory the 14 identity and spatial locations of multiple talkers and inhibiting the representations of irrelevant 15 talkers. The experiments of Beer and colleagues differ from the current experiments in that they used 16 parent reports of executive function abilities (Beer et al., 2011) and visual tests of executive function 17 (Beer et al., 2014). Therefore, a comparison between the current experiment and the experiments of 18 Beer and colleagues does not reveal whether the types or extent of executive function deficits differ 19 between children with moderate and children with severe-to-profound hearing loss. 20 Children with severe-to-profound hearing loss might be expected to show greater deficits in 21 executive function abilities, or perhaps a wider variety of executive function abilities that are affected, 22 than children with moderate hearing loss. That prediction follows from the idea that children with 23 severe-to-profound hearing loss would have experienced a period of time (between the onset of 24 hearing loss and receiving cochlear implants) during which they were more deprived of acoustic 25 stimulation than children with moderate hearing losses (who would have experienced a delay 26 between the onset of hearing loss and receiving hearing aids, but who have greater preservation of 23
24 residual hearing). In addition, CIs and hearing aids provide different types of acoustic information to 2 the listener that may affect the ability of executive functions to develop after rehabilitation. The 3 current experiment reveals that children with moderate hearing loss show atypical preparatory 4 attention during multi-talker listening, which might relate directly to the difficulty they experience in 5 multi-talker environments; however, it does not reveal whether other executive functions, including 6 those in other sensory modalities, are atypical. Nevertheless, a link between the lack of preparatory 7 activity obtained in the current experiment and broader executive function abilities is possible 8 because the development of executive functions, such as working memory, has been related to the 9 strength of the CNV (Segalowitz and Davies, 2004). Greater understanding of how hearing loss affects 10 executive function could be gained by directly comparing individuals with different hearing loss 11 aetiologies on the same executive function tasks. In addition, it would be informative for future studies 12 to examine the relationship between preparatory attention during multi-talker listening and a broader 13 range of executive function abilities Implications 15 Current interventions for impaired hearing, such as acoustic hearing aids, are targeted at 16 overcoming a loss of sensitivity at the auditory periphery. The current results have potential 17 implications for rehabilitation, because they suggest that atypical auditory attention might be one 18 factor that contributes to difficulty understanding speech for HI children during multi-talker listening. 19 Although it is currently unclear how attention abilities could be restored, improving auditory attention 20 abilities (e.g. through training) might help hearing-impaired children to understand speech in the 21 presence of other competing speech a situation that would frequently be encountered in noisy 22 environments at home and at school. 23 Better understanding of the conditions under which hearing loss affects attention and the 24 extent to which hearing loss affects other executive functions is required to identify the underlying 25 cause of atypical attention in hearing-impaired children. This knowledge may provide insights into 26 novel strategies by which auditory attention could be restored in hearing-impaired children. If 24
25 directing preparatory attention relies on accurate representations of the cues used to direct attention, 2 focusing on improving those cues may be desirable for future rehabilitation. Whereas, if a wider 3 variety of executive functions are affected by hearing loss, then cognitive training may be more 4 appropriate (see Posner et al., 2015, for a review). The success of these rehabilitation techniques may 5 also depend on whether a critical or sensitive period exists for the development of executive functions. 6 Given there may be individual variability in executive function ability depending on the extent of 7 hearing loss or age of onset, different rehabilitation strategies may be best suited to different 8 individuals. Future experiments should aim to identify whether hearing loss aetiology affects 9 executive function and whether it is possible to restore preparatory brain activity in hearing-impaired 10 children Conclusion 12 The results demonstrate that moderate sensorineural hearing loss has consequences for 13 central auditory processing. When presented with a visual cue that directed attention to the location 14 of an upcoming talker, NH children utilised preparatory brain activity. The group of HI children showed 15 significantly weaker evidence of preparatory brain activity than the group of NH children. This result 16 suggests that, on average, HI children do not direct preparatory spatial attention to the same extent 17 as NH children of a similar age. In addition, preparatory spatial attention was not restored when HI 18 children listened using their acoustic hearing aids. Consequently, difficulties with preparatory 19 attention in hearing-impaired children are likely to contribute to difficulties understanding speech in 20 noisy acoustic environments. 25
26 Acknowledgements 2 This work was supported by a studentship from the Goodricke Appeal Fund to EH. For their 3 help recruiting patients, we thank Gerard Reilly and Kate Iley of the York Teaching Hospital NHS 4 Foundation Trust, Aung Nyunt of Hull and East Yorkshire Hospitals NHS Trust, Sanjay Verma and 5 Mirriam Iqbal of The Leeds Teaching Hospitals NHS Trust, and Christopher Raine, Rob Gardner, and 6 Sara Morgan of Bradford Teaching Hospitals NHS Foundation Trust. 26
27 References 2 Alain, C., Roye, A., Salloum, C., Effects of age-related hearing loss and background noise on 3 neuromagnetic activity from auditory cortex. Front. Syst. Neurosci. 8, 8. 4 doi: /fnsys Assmann, P.F., Summerfield, A.Q., The contribution of waveform interactions to the 6 perception of concurrent vowels. J. Acoust. Soc. Am. 95, Banerjee, S., Snyder, A.C., Molholm, S., Foxe, J.J., Oscillatory alpha-band mechanisms and the 8 deployment of spatial attention to anticipated auditory and visual target locations: Supramodal 9 or sensory-specific control. J. Neurosci. 31, doi: /jneurosci Oscillatory 11 Basile, L.F., Brunder, D.G., Tarkka, I.M., Papanicolaou, A.C., Magnetic fields from human 12 prefrontal cortex differ during two recognition tasks. Int. J. Psychophysiol. 27, Beer, J., Kronenberger, W.G., Castellanos, I., Colson, B.G., Henning, S.C., Pisoni, D.B., Executive 14 Functioning Skills in Preschool-Age Children With Cochlear Implants. J. Speech, Lang. Hear. Res , doi: / Beer, J., Kronenberger, W.G., Pisoni, D.B., Executive function in everyday life: implications for 17 young cochlear implant users. Cochlear Implants Int. 12, S doi: /j.pestbp investigations 19 Best, V., Marrone, N., Mason, C.R., Kidd, G., Shinn-Cunningham, B.G., Effects of sensorineural 20 hearing loss on visually guided attention in a multitalker environment. J. Assoc. Res. 21 Otolaryngol. 10, doi: /s Best, V., Ozmeral, E.J., Shinn-Cunningham, B.G., Visually-guided attention enhances target 23 identification in a complex auditory scene. J. Assoc. Res. Otolaryngol. 8, doi: /s z 27
28 Bronkhorst, A.W., Plomp, R., The effect of head-induced interaural time and level differences 2 on speech intelligibility in noise. J. Acoust. Soc. Am. 83, Brungart, D.S., Informational and energetic masking effects in the perception of two 4 simultaneous talkers. J. Acoust. Soc. Am. 109, doi: / Brungart, D.S., Simpson, B.D., Ericson, M.A., Scott, K.R., Informational and energetic masking 6 effects in the perception of multiple simultaneous talkers. J. Acoust. Soc. Am. 110, doi: / Campbell, J., Sharma, A., Compensatory changes in cortical resource allocation in adults with 9 hearing loss. Front. Syst. Neurosci. 7, 71. doi: /fnsys Cardin, V., Effects of aging and adult-onset hearing loss on cortical auditory regions. Front. 11 Neurosci. 10, doi: /fnins Chennu, S., Noreika, V., Gueorguiev, D., Blenkmann, A., Kochen, S., Ibáñez, A., Owen, A.M., 13 Bekinschtein, T.A., Expectation and attention in hierarchical auditory prediction. J. 14 Neurosci. 33, doi: /jneurosci Darwin, C.J., Hukin, R.W., Effectiveness of spatial cues, prosody, and talker characteristics in 16 selective attention. J. Acoust. Soc. Am. 107, Darwin, C.J., Hukin, R.W., Auditory objects of attention: the role of interaural time differences. 18 J. Exp. Psychol. Hum. Percept. Perform. 25, Dubno, J.R., Dirks, D.D., Morgan, D.E., Effects of age and mild hearing loss on speech 20 recognition in noise. J. Acoust. Soc. Am. 76, Ericson, M.A., Brungart, D.S., Brian, D., Factors that influence intelligibility in multitalker 22 speech displays. Int. J. Aviat. Psychol. 14, Freyman, R.L., Balakrishnan, U., Helfer, K.S., Effect of number of masking talkers and auditory 24 priming on informational masking in speech recognition. J. Acoust. Soc. Am. 115,
29 doi: / Gaudrain, E., Grimault, N., Healy, E.W., Béra, J., Effect of spectral smearing on the perceptual 3 segregation of vowel sequences. Hear. Res. 231, Helfer, K.S., Freyman, R.L., Aging and speech-on-speech masking. Ear Hear. 29, doi: /aud.0b013e31815d638b.aging 6 Helfer, K.S., Freyman, R.L., The role of visual speech cues in reducing energetic and 7 informational masking. J. Acoust. Soc. Am. 117, doi: / Hilbert, D., Grundzüge einer Allgemeinen Theorie der linearen Integralgleichungen (Basics of a 9 general theory of linear integral equations). Teubner, Leizpig. 10 Hill, K.T., Miller, L.M., Auditory attentional control and selection during cocktail party listening. 11 Cereb. Cortex 20, doi: /cercor/bhp Hogan, C.A., Turner, C.W., High-frequency audibility: Benefits for hearing-impaired listeners. J. 13 Acoust. Soc. Am. 104, doi: / Holmes, E., Kitterick, P.T., Summerfield, A.Q., EEG activity evoked in preparation for multi- 15 talker listening by adults and children. Hear. Res. 336, doi: /j.heares Kidd, G., Arbogast, T.L., Mason, C.R., Gallun, F.J., The advantage of knowing where to listen. J. 18 Acoust. Soc. Am. 118, doi: / Kitterick, P.T., Bailey, P.J., Summerfield, A.Q., Benefits of knowing who, where, and when in 20 multi-talker listening. J. Acoust. Soc. Am. 127, doi: / Koravand, A., Jutras, B., Lassonde, M., Cortical auditory evoked potentials in children with a 22 hearing loss: a pilot study. Int. J. Pediatr. 2012, doi: /2012/ Larson, E., Lee, A.K.C., Switching auditory attention using spatial and non-spatial features 24 recruits different cortical networks. Neuroimage 84,
30 doi: /j.neuroimage Lee, A.K.C., Rajaram, S., Xia, J., Bharadwaj, H., Larson, E., Hämäläinen, M.S., Shinn-Cunningham, B.G., Auditory selective attention reveals preparatory activity in different cortical regions for 4 selection based on source location and source pitch. Front. Neurosci. 6, doi: /fnins Lorenzi, C., Gilbert, G., Carn, H., Garnier, S., Moore, B.C.J., Speech perception problems of the 7 hearing impaired reflect inability to use temporal fine structure. Proc. Natl. Acad. Sci. U. S. A , doi: /pnas Maris, E., Oostenveld, R., Nonparametric statistical testing of EEG- and MEG-data. J. Neurosci. 10 Methods 164, doi: /j.jneumeth Moore, B.C.J., The role of temporal fine structure processing in pitch perception, masking, and 12 speech perception for normal-hearing and hearing-impaired people. J. Assoc. Res. Otolaryngol. 13 9, doi: /s x 14 Moore, B.C.J., Cochlear Hearing Loss. Whurr Publishers Ltd., London. 15 Moore, T.J., Voice communication jamming research, in: AGARD Conference Proceedings 331: 16 Aural Communication in Aviation. Neuilly-Sur-Seine, France, p. 2:1-2:6. 17 Müller, N., Weisz, N., Lateralized auditory cortical alpha band activity and interregional 18 connectivity pattern reflect anticipation of target sounds. Cereb. Cortex 22, doi: /cercor/bhr Neher, T., Behrens, T., Carlile, S., Jin, C., Kragelund, L., Petersen, A.S., Schaik, A. Van, Benefit 21 from spatial separation of multiple talkers in bilateral hearing-aid users: Effects of hearing loss, 22 age, and cognition. Int. J. Audiol. 48, doi: / O Sullivan, J.A., Power, A.J., Mesgarani, N., Rajaram, S., Foxe, J.J., Shinn-Cunningham, B.G., Slaney, 24 M., Shamma, S.A., Lalor, E.C., Attentional selection in a cocktail party environment can 30
31 be decoded from single-trial EEG. Cereb. Cortex doi: /cercor/bht355 2 Oates, P.A., Kurtzberg, D., Stapells, D.R., Effects of Sensorineural Hearing Loss on Cortical 3 Event-Related Potential and Behavioral Measures of Speech-Sound Processing. Ear Hear. 23, doi: /01.aud Pasinski, A.C., Mcauley, J.D., Snyder, J.S., How modality specific is processing of auditory and 6 visual rhythms? Psychophysiology 53, doi: /psyp Peelle, J.E., Troiani, V., Grossman, M., Wingfield, A., Hearing loss in older adults affects neural 8 systems supporting speech comprehension. J. Neurosci. 31, doi: /jneurosci Posner, M.I., Rothbart, M.K., Tang, Y.Y., Enhancing attention through training. Curr. Opin. 11 Behav. Sci. 4, 1 5. doi: /j.cobeha Robertson, I.H., Ward, T., Ridgeway, V., The structure of normal human attention: The Test of 13 Everyday Attention. J. Int. Neuropsychol. Soc. 2, doi: /s Rockstroh, B., Müller, M., Wagner, M., Cohen, R., Elbert, T., Probing the nature of the CNV. 15 Electroencephalogr. Clin. Neurophysiol. 87, doi: / (93)90023-o 16 Ruchkin, D.S., McCalley, M.G., Glaser, E.M., Event Related Potentials and Time Estimation. 17 Psychophysiology. doi: /j tb01311.x 18 Segalowitz, S.J., Davies, P.L., Charting the maturation of the frontal lobe: An 19 electrophysiological strategy. Brain Cogn. 55, doi: /s (03) Shafiro, V., Gygi, B., Perceiving the speech of multiple concurrent talkers in a combined 21 divided and selective attention task. J. Acoust. Soc. Am. 122, EL doi: / Tecce, J.J., Scheff, N.M., Attention Reduction and Suppressed Direct-Current Potentials in the 23 Human Brain. Science (80). 164, doi: / Travis, F., Tecce, J.J., Effects of distracting stimuli on CNV amplitude and reaction time. Int. J. 31
32 Psychophysiol. 31, doi: /s (98) Voisin, J., Bidet-Caulet, A., Bertrand, O., Fonlupt, P., Listening in silence activates auditory 3 areas: a functional magnetic resonance imaging study. J. Neurosci. 26, doi: /jneurosci Walter, W.G., Cooper, R., Aldridge, V.J., McCallum, W.C., Winter, A.L., Contingent negative 6 variation: An electric sign of sensori-motor association and expectancy in the human brain. 7 Nature 203, doi: /203380a0 8 Wöstmann, M., Schröger, E., Obleser, J., Acoustic detail guides attention allocation in a 9 selective listening task. J. Cogn. Neurosci. 27, doi: /jocn 32
33 Figure Captions 2 3 Fig. 1. Average pure-tone audiometric thresholds (db HL) for hearing-impaired (HI; N = 14) and 4 normally-hearing (NH; N = 24) children, plotted separately for the left (A) and right (B) ears. Grey 5 dashed lines show thresholds for individual hearing-impaired participants and the black solid lines 6 show mean thresholds across HI (diamonds) and NH (circles) participants. 7 8 Fig. 2. (A) Layout of loudspeakers (dark grey squares) and visual display unit (light grey rectangle) 9 relative to a participant's head. Visual cues for location (B,C) and gender (D,E). A visual composite 10 stimulus (F) was created by overlaying the four visual cues Fig. 3. Schematic showing the trial structure in the test condition (A) and the control condition (B). 13 Stimuli for an example trial are displayed below, with an example of the visual stimuli (left; attend- 14 left trial), acoustical stimuli (centre) and response buttons (right) Fig. 4. Schematic of EEG analysis pipeline. An example is provided for the comparison between the 17 test and control conditions. (A) EEG data were pre-processed and averaged across trials, producing 18 time-locked event-related potentials (ERPs) at each electrode for each participant. (B) Spatio- 19 temporal cluster-based permutation analysis was used to extract clusters of electrodes and time 20 points that differed significantly between conditions. An example is shown, in which the scalp map 21 shows the electrodes that contributed to the cluster (red circles), the graph illustrates ERPs at those 22 electrodes, and the dashed box on the graph indicates the time window of each cluster. Time on the 23 x-axis is relative to the onset of the visual cues. (C) For each cluster, a bootstrapped null distribution 24 was assembled by selecting, with replacement, samples of NH children of equal size to the 33
34 comparison group of HI children. For each sample, the average cluster magnitude was calculated as 2 the difference in amplitude between conditions, averaged across the electrodes and time points that 3 contributed to the cluster. (D) The average cluster magnitude in HI children was compared to the 4 bootstrapped distribution from NH children in a two-tailed test. 5 6 Fig. 5. Mean percentage of trials in which participants correctly identified the colour-number 7 combination spoken by the target talker in the test condition. Separate bars illustrate the results for 8 normally-hearing children (NH; N = 24), hearing-impaired children listening unaided (HI U ; N = 14), 9 and hearing-impaired children listening aided (HI U ; N = 10). Error bars show ±1 standard error of the 10 mean Fig. 6. Results from Spatio-temporal cluster-based permutation analyses in normally-hearing (NH) 13 children for Location (A D) and Gender (E F) trials. (A and E) Coloured rectangles indicate the time- 14 span of significant (p < 0.05) clusters of activity. Time on the x-axis is relative to the onset of the 15 visual cues. Rows on the y-axis show separate significant clusters. For clusters plotted as red 16 rectangles, the average amplitude, over all space-by-time points in the cluster, was more positive in 17 the test condition than the control condition. For clusters plotted as blue rectangles, the average 18 amplitude was more negative in the test condition than the control condition. Further information 19 about each cluster is displayed in (B D and F). For each cluster, the topographical map shows the 20 average topography across the time-span of the cluster and black circles superimposed on the 21 topographical map show electrodes that contributed to the cluster. The graph shows ERPs averaged 22 across the electrodes that contributed to the cluster and the dashed grey rectangle indicates the 23 time-span of the cluster Fig. 7. Cluster size differed between normally-hearing (NH; N = 24) and hearing-impaired children 34
35 (HI U ; N = 14) for the clusters that occurred during location trials (A), but not for the cluster that 2 occurred during gender trials (B). For Clusters 2N and 2P, we observed similar results when 3 comparing NH children with the sub-set of hearing-impaired children who completed the task with 4 their hearing aids (HI A ; N = 10). Error bars for HI U and HI A children show 95% confidence intervals for 5 each group. Error bars for NH children show 95% confidence intervals from the bootstrapped null 6 distribution. Brackets above each cluster indicate whether there was a significant difference 7 between the groups (* p < 0.050; ** p < 0.010; *** p < 0.001; n.s. not significant). The time window 8 of the cluster and the electrodes which contributed are displayed above each cluster Fig. 8. Results from Spatio-temporal cluster-based permutation analyses in hearing-impaired 11 children (listening unaided; HI U group) for Location (A) and Gender (B C) trials. (A and B) Coloured 12 rectangles indicate the time-span of significant (p < 0.05) clusters of activity. Time on the x-axis is 13 relative to the onset of the visual cues. Rows on the y-axis show separate significant clusters. No 14 significant clusters were found for location trials. For clusters plotted as blue rectangles, the average 15 amplitude was more negative in the test condition than the control condition. Further information 16 about each cluster is displayed in (C). The topographical map shows the average topography across 17 the time-span of the cluster and black circles superimposed on the topographical map show 18 electrodes that contributed to the cluster. The graph shows ERPs averaged across the electrodes 19 that contributed to the cluster and the dashed grey rectangle indicates the time-span of the cluster. 20 (D) Cluster size did not differ signfiicantly between normally-hearing (NH; N = 24) and hearing- 21 impaired children (HI U ; N = 14) for the cluster that occurred during gender trials. The error bar for 22 HI U children shows the 95% confidence interval. The error bar for NH children shows the 95% 23 confidence interval from the bootstrapped null distribution. 35
36
37
38
39
40
DAT335 Music Perception and Cognition Cogswell Polytechnical College Spring Week 6 Class Notes
 DAT335 Music Perception and Cognition Cogswell Polytechnical College Spring 2009 Week 6 Class Notes Pitch Perception Introduction Pitch may be described as that attribute of auditory sensation in terms
DAT335 Music Perception and Cognition Cogswell Polytechnical College Spring 2009 Week 6 Class Notes Pitch Perception Introduction Pitch may be described as that attribute of auditory sensation in terms
HBI Database. Version 2 (User Manual)
") HBI Database Version 2 (User Manual) St-Petersburg, Russia 2007 2 1. INTRODUCTION...3 2. RECORDING CONDITIONS...6 2.1. EYE OPENED AND EYE CLOSED CONDITION....6 2.2. VISUAL CONTINUOUS PERFORMANCE TASK...6
HBI Database Version 2 (User Manual) St-Petersburg, Russia 2007 2 1. INTRODUCTION...3 2. RECORDING CONDITIONS...6 2.1. EYE OPENED AND EYE CLOSED CONDITION....6 2.2. VISUAL CONTINUOUS PERFORMANCE TASK...6
I. INTRODUCTION. Electronic mail:
 Neural activity associated with distinguishing concurrent auditory objects Claude Alain, a) Benjamin M. Schuler, and Kelly L. McDonald Rotman Research Institute, Baycrest Centre for Geriatric Care, 3560
Neural activity associated with distinguishing concurrent auditory objects Claude Alain, a) Benjamin M. Schuler, and Kelly L. McDonald Rotman Research Institute, Baycrest Centre for Geriatric Care, 3560
Music Perception with Combined Stimulation
 Music Perception with Combined Stimulation Kate Gfeller 1,2,4, Virginia Driscoll, 4 Jacob Oleson, 3 Christopher Turner, 2,4 Stephanie Kliethermes, 3 Bruce Gantz 4 School of Music, 1 Department of Communication
Music Perception with Combined Stimulation Kate Gfeller 1,2,4, Virginia Driscoll, 4 Jacob Oleson, 3 Christopher Turner, 2,4 Stephanie Kliethermes, 3 Bruce Gantz 4 School of Music, 1 Department of Communication
Pre-Processing of ERP Data. Peter J. Molfese, Ph.D. Yale University
 Pre-Processing of ERP Data Peter J. Molfese, Ph.D. Yale University Before Statistical Analyses, Pre-Process the ERP data Planning Analyses Waveform Tools Types of Tools Filter Segmentation Visual Review
Pre-Processing of ERP Data Peter J. Molfese, Ph.D. Yale University Before Statistical Analyses, Pre-Process the ERP data Planning Analyses Waveform Tools Types of Tools Filter Segmentation Visual Review
Behavioral and neural identification of birdsong under several masking conditions
 Behavioral and neural identification of birdsong under several masking conditions Barbara G. Shinn-Cunningham 1, Virginia Best 1, Micheal L. Dent 2, Frederick J. Gallun 1, Elizabeth M. McClaine 2, Rajiv
Behavioral and neural identification of birdsong under several masking conditions Barbara G. Shinn-Cunningham 1, Virginia Best 1, Micheal L. Dent 2, Frederick J. Gallun 1, Elizabeth M. McClaine 2, Rajiv
DATA! NOW WHAT? Preparing your ERP data for analysis
 DATA! NOW WHAT? Preparing your ERP data for analysis Dennis L. Molfese, Ph.D. Caitlin M. Hudac, B.A. Developmental Brain Lab University of Nebraska-Lincoln 1 Agenda Pre-processing Preparing for analysis
DATA! NOW WHAT? Preparing your ERP data for analysis Dennis L. Molfese, Ph.D. Caitlin M. Hudac, B.A. Developmental Brain Lab University of Nebraska-Lincoln 1 Agenda Pre-processing Preparing for analysis
AUD 6306 Speech Science
 AUD 3 Speech Science Dr. Peter Assmann Spring semester 2 Role of Pitch Information Pitch contour is the primary cue for tone recognition Tonal languages rely on pitch level and differences to convey lexical
AUD 3 Speech Science Dr. Peter Assmann Spring semester 2 Role of Pitch Information Pitch contour is the primary cue for tone recognition Tonal languages rely on pitch level and differences to convey lexical
Pitch is one of the most common terms used to describe sound.
 ARTICLES https://doi.org/1.138/s41562-17-261-8 Diversity in pitch perception revealed by task dependence Malinda J. McPherson 1,2 * and Josh H. McDermott 1,2 Pitch conveys critical information in speech,
ARTICLES https://doi.org/1.138/s41562-17-261-8 Diversity in pitch perception revealed by task dependence Malinda J. McPherson 1,2 * and Josh H. McDermott 1,2 Pitch conveys critical information in speech,
Precedence-based speech segregation in a virtual auditory environment
 Precedence-based speech segregation in a virtual auditory environment Douglas S. Brungart a and Brian D. Simpson Air Force Research Laboratory, Wright-Patterson AFB, Ohio 45433 Richard L. Freyman University
Precedence-based speech segregation in a virtual auditory environment Douglas S. Brungart a and Brian D. Simpson Air Force Research Laboratory, Wright-Patterson AFB, Ohio 45433 Richard L. Freyman University
Measurement of overtone frequencies of a toy piano and perception of its pitch
 Measurement of overtone frequencies of a toy piano and perception of its pitch PACS: 43.75.Mn ABSTRACT Akira Nishimura Department of Media and Cultural Studies, Tokyo University of Information Sciences,
Measurement of overtone frequencies of a toy piano and perception of its pitch PACS: 43.75.Mn ABSTRACT Akira Nishimura Department of Media and Cultural Studies, Tokyo University of Information Sciences,
Voice segregation by difference in fundamental frequency: Effect of masker type
 Voice segregation by difference in fundamental frequency: Effect of masker type Mickael L. D. Deroche a) Department of Otolaryngology, Johns Hopkins University School of Medicine, 818 Ross Research Building,
Voice segregation by difference in fundamental frequency: Effect of masker type Mickael L. D. Deroche a) Department of Otolaryngology, Johns Hopkins University School of Medicine, 818 Ross Research Building,
User Guide Slow Cortical Potentials (SCP)
") User Guide Slow Cortical Potentials (SCP) This user guide has been created to educate and inform the reader about the SCP neurofeedback training protocol for the NeXus 10 and NeXus-32 systems with the
User Guide Slow Cortical Potentials (SCP) This user guide has been created to educate and inform the reader about the SCP neurofeedback training protocol for the NeXus 10 and NeXus-32 systems with the
Object selectivity of local field potentials and spikes in the macaque inferior temporal cortex
 Object selectivity of local field potentials and spikes in the macaque inferior temporal cortex Gabriel Kreiman 1,2,3,4*#, Chou P. Hung 1,2,4*, Alexander Kraskov 5, Rodrigo Quian Quiroga 6, Tomaso Poggio
Object selectivity of local field potentials and spikes in the macaque inferior temporal cortex Gabriel Kreiman 1,2,3,4*#, Chou P. Hung 1,2,4*, Alexander Kraskov 5, Rodrigo Quian Quiroga 6, Tomaso Poggio
PROCESSING YOUR EEG DATA
 PROCESSING YOUR EEG DATA Step 1: Open your CNT file in neuroscan and mark bad segments using the marking tool (little cube) as mentioned in class. Mark any bad channels using hide skip and bad. Save the
PROCESSING YOUR EEG DATA Step 1: Open your CNT file in neuroscan and mark bad segments using the marking tool (little cube) as mentioned in class. Mark any bad channels using hide skip and bad. Save the
Preparation of the participant. EOG, ECG, HPI coils : what, why and how
 Preparation of the participant EOG, ECG, HPI coils : what, why and how 1 Introduction In this module you will learn why EEG, ECG and HPI coils are important and how to attach them to the participant. The
Preparation of the participant EOG, ECG, HPI coils : what, why and how 1 Introduction In this module you will learn why EEG, ECG and HPI coils are important and how to attach them to the participant. The
Pitch. The perceptual correlate of frequency: the perceptual dimension along which sounds can be ordered from low to high.
 Pitch The perceptual correlate of frequency: the perceptual dimension along which sounds can be ordered from low to high. 1 The bottom line Pitch perception involves the integration of spectral (place)
Pitch The perceptual correlate of frequency: the perceptual dimension along which sounds can be ordered from low to high. 1 The bottom line Pitch perception involves the integration of spectral (place)
Acoustic and musical foundations of the speech/song illusion
 Acoustic and musical foundations of the speech/song illusion Adam Tierney, *1 Aniruddh Patel #2, Mara Breen^3 * Department of Psychological Sciences, Birkbeck, University of London, United Kingdom # Department
Acoustic and musical foundations of the speech/song illusion Adam Tierney, *1 Aniruddh Patel #2, Mara Breen^3 * Department of Psychological Sciences, Birkbeck, University of London, United Kingdom # Department
Pitch Perception and Grouping. HST.723 Neural Coding and Perception of Sound
 Pitch Perception and Grouping HST.723 Neural Coding and Perception of Sound Pitch Perception. I. Pure Tones The pitch of a pure tone is strongly related to the tone s frequency, although there are small
Pitch Perception and Grouping HST.723 Neural Coding and Perception of Sound Pitch Perception. I. Pure Tones The pitch of a pure tone is strongly related to the tone s frequency, although there are small
Proceedings of Meetings on Acoustics
 Proceedings of Meetings on Acoustics Volume 19, 2013 http://acousticalsociety.org/ ICA 2013 Montreal Montreal, Canada 2-7 June 2013 Psychological and Physiological Acoustics Session 1pPPb: Psychoacoustics
Proceedings of Meetings on Acoustics Volume 19, 2013 http://acousticalsociety.org/ ICA 2013 Montreal Montreal, Canada 2-7 June 2013 Psychological and Physiological Acoustics Session 1pPPb: Psychoacoustics
Untangling syntactic and sensory processing: An ERP study of music perception
 Manuscript accepted for publication in Psychophysiology Untangling syntactic and sensory processing: An ERP study of music perception Stefan Koelsch, Sebastian Jentschke, Daniela Sammler, & Daniel Mietchen
Manuscript accepted for publication in Psychophysiology Untangling syntactic and sensory processing: An ERP study of music perception Stefan Koelsch, Sebastian Jentschke, Daniela Sammler, & Daniel Mietchen
Common Spatial Patterns 3 class BCI V Copyright 2012 g.tec medical engineering GmbH
 g.tec medical engineering GmbH Sierningstrasse 14, A-4521 Schiedlberg Austria - Europe Tel.: (43)-7251-22240-0 Fax: (43)-7251-22240-39 office@gtec.at, http://www.gtec.at Common Spatial Patterns 3 class
g.tec medical engineering GmbH Sierningstrasse 14, A-4521 Schiedlberg Austria - Europe Tel.: (43)-7251-22240-0 Fax: (43)-7251-22240-39 office@gtec.at, http://www.gtec.at Common Spatial Patterns 3 class
The Influence of Explicit Markers on Slow Cortical Potentials During Figurative Language Processing
 The Influence of Explicit Markers on Slow Cortical Potentials During Figurative Language Processing Christopher A. Schwint (schw6620@wlu.ca) Department of Psychology, Wilfrid Laurier University 75 University
The Influence of Explicit Markers on Slow Cortical Potentials During Figurative Language Processing Christopher A. Schwint (schw6620@wlu.ca) Department of Psychology, Wilfrid Laurier University 75 University
Non-native Homonym Processing: an ERP Measurement
 Non-native Homonym Processing: an ERP Measurement Jiehui Hu ab, Wenpeng Zhang a, Chen Zhao a, Weiyi Ma ab, Yongxiu Lai b, Dezhong Yao b a School of Foreign Languages, University of Electronic Science &
Non-native Homonym Processing: an ERP Measurement Jiehui Hu ab, Wenpeng Zhang a, Chen Zhao a, Weiyi Ma ab, Yongxiu Lai b, Dezhong Yao b a School of Foreign Languages, University of Electronic Science &
Brain-Computer Interface (BCI)
") Brain-Computer Interface (BCI) Christoph Guger, Günter Edlinger, g.tec Guger Technologies OEG Herbersteinstr. 60, 8020 Graz, Austria, guger@gtec.at This tutorial shows HOW-TO find and extract proper signal
Brain-Computer Interface (BCI) Christoph Guger, Günter Edlinger, g.tec Guger Technologies OEG Herbersteinstr. 60, 8020 Graz, Austria, guger@gtec.at This tutorial shows HOW-TO find and extract proper signal
The N400 and Late Positive Complex (LPC) Effects Reflect Controlled Rather than Automatic Mechanisms of Sentence Processing
 Effects Reflect Controlled Rather than Automatic Mechanisms of Sentence Processing") Brain Sci. 2012, 2, 267-297; doi:10.3390/brainsci2030267 Article OPEN ACCESS brain sciences ISSN 2076-3425 www.mdpi.com/journal/brainsci/ The N400 and Late Positive Complex (LPC) Effects Reflect Controlled
Brain Sci. 2012, 2, 267-297; doi:10.3390/brainsci2030267 Article OPEN ACCESS brain sciences ISSN 2076-3425 www.mdpi.com/journal/brainsci/ The N400 and Late Positive Complex (LPC) Effects Reflect Controlled
Common Spatial Patterns 2 class BCI V Copyright 2012 g.tec medical engineering GmbH
 g.tec medical engineering GmbH Sierningstrasse 14, A-4521 Schiedlberg Austria - Europe Tel.: (43)-7251-22240-0 Fax: (43)-7251-22240-39 office@gtec.at, http://www.gtec.at Common Spatial Patterns 2 class
g.tec medical engineering GmbH Sierningstrasse 14, A-4521 Schiedlberg Austria - Europe Tel.: (43)-7251-22240-0 Fax: (43)-7251-22240-39 office@gtec.at, http://www.gtec.at Common Spatial Patterns 2 class
Temporal Envelope and Periodicity Cues on Musical Pitch Discrimination with Acoustic Simulation of Cochlear Implant
 Temporal Envelope and Periodicity Cues on Musical Pitch Discrimination with Acoustic Simulation of Cochlear Implant Lichuan Ping 1, 2, Meng Yuan 1, Qinglin Meng 1, 2 and Haihong Feng 1 1 Shanghai Acoustics
Temporal Envelope and Periodicity Cues on Musical Pitch Discrimination with Acoustic Simulation of Cochlear Implant Lichuan Ping 1, 2, Meng Yuan 1, Qinglin Meng 1, 2 and Haihong Feng 1 1 Shanghai Acoustics
How Order of Label Presentation Impacts Semantic Processing: an ERP Study
 How Order of Label Presentation Impacts Semantic Processing: an ERP Study Jelena Batinić (jelenabatinic1@gmail.com) Laboratory for Neurocognition and Applied Cognition, Department of Psychology, Faculty
How Order of Label Presentation Impacts Semantic Processing: an ERP Study Jelena Batinić (jelenabatinic1@gmail.com) Laboratory for Neurocognition and Applied Cognition, Department of Psychology, Faculty
MEASURING LOUDNESS OF LONG AND SHORT TONES USING MAGNITUDE ESTIMATION
 MEASURING LOUDNESS OF LONG AND SHORT TONES USING MAGNITUDE ESTIMATION Michael Epstein 1,2, Mary Florentine 1,3, and Søren Buus 1,2 1Institute for Hearing, Speech, and Language 2Communications and Digital
MEASURING LOUDNESS OF LONG AND SHORT TONES USING MAGNITUDE ESTIMATION Michael Epstein 1,2, Mary Florentine 1,3, and Søren Buus 1,2 1Institute for Hearing, Speech, and Language 2Communications and Digital
A 5 Hz limit for the detection of temporal synchrony in vision
 A 5 Hz limit for the detection of temporal synchrony in vision Michael Morgan 1 (Applied Vision Research Centre, The City University, London) Eric Castet 2 ( CRNC, CNRS, Marseille) 1 Corresponding Author
A 5 Hz limit for the detection of temporal synchrony in vision Michael Morgan 1 (Applied Vision Research Centre, The City University, London) Eric Castet 2 ( CRNC, CNRS, Marseille) 1 Corresponding Author
Music BCI ( )
") Music BCI (006-2015) Matthias Treder, Benjamin Blankertz Technische Universität Berlin, Berlin, Germany September 5, 2016 1 Introduction We investigated the suitability of musical stimuli for use in a
Music BCI (006-2015) Matthias Treder, Benjamin Blankertz Technische Universität Berlin, Berlin, Germany September 5, 2016 1 Introduction We investigated the suitability of musical stimuli for use in a
Release from speech-on-speech masking in a front-and-back geometry
 Release from speech-on-speech masking in a front-and-back geometry Neil L. Aaronson Department of Physics and Astronomy, Michigan State University, Biomedical and Physical Sciences Building, East Lansing,
Release from speech-on-speech masking in a front-and-back geometry Neil L. Aaronson Department of Physics and Astronomy, Michigan State University, Biomedical and Physical Sciences Building, East Lansing,
MEANING RELATEDNESS IN POLYSEMOUS AND HOMONYMOUS WORDS: AN ERP STUDY IN RUSSIAN
 Anna Yurchenko, Anastasiya Lopukhina, Olga Dragoy MEANING RELATEDNESS IN POLYSEMOUS AND HOMONYMOUS WORDS: AN ERP STUDY IN RUSSIAN BASIC RESEARCH PROGRAM WORKING PAPERS SERIES: LINGUISTICS WP BRP 67/LNG/2018
Anna Yurchenko, Anastasiya Lopukhina, Olga Dragoy MEANING RELATEDNESS IN POLYSEMOUS AND HOMONYMOUS WORDS: AN ERP STUDY IN RUSSIAN BASIC RESEARCH PROGRAM WORKING PAPERS SERIES: LINGUISTICS WP BRP 67/LNG/2018
23/01/51. Gender-selective effects of the P300 and N400 components of the. VEP waveform. How are ERP related to gender? Event-Related Potential (ERP)
") 23/01/51 EventRelated Potential (ERP) Genderselective effects of the and N400 components of the visual evoked potential measuring brain s electrical activity (EEG) responded to external stimuli EEG averaging
23/01/51 EventRelated Potential (ERP) Genderselective effects of the and N400 components of the visual evoked potential measuring brain s electrical activity (EEG) responded to external stimuli EEG averaging
Consonance perception of complex-tone dyads and chords
 Downloaded from orbit.dtu.dk on: Nov 24, 28 Consonance perception of complex-tone dyads and chords Rasmussen, Marc; Santurette, Sébastien; MacDonald, Ewen Published in: Proceedings of Forum Acusticum Publication
Downloaded from orbit.dtu.dk on: Nov 24, 28 Consonance perception of complex-tone dyads and chords Rasmussen, Marc; Santurette, Sébastien; MacDonald, Ewen Published in: Proceedings of Forum Acusticum Publication
The Tone Height of Multiharmonic Sounds. Introduction
 Music-Perception Winter 1990, Vol. 8, No. 2, 203-214 I990 BY THE REGENTS OF THE UNIVERSITY OF CALIFORNIA The Tone Height of Multiharmonic Sounds ROY D. PATTERSON MRC Applied Psychology Unit, Cambridge,
Music-Perception Winter 1990, Vol. 8, No. 2, 203-214 I990 BY THE REGENTS OF THE UNIVERSITY OF CALIFORNIA The Tone Height of Multiharmonic Sounds ROY D. PATTERSON MRC Applied Psychology Unit, Cambridge,
ARTICLE IN PRESS BRESC-40606; No. of pages: 18; 4C:
 BRESC-40606; No. of pages: 18; 4C: DTD 5 Cognitive Brain Research xx (2005) xxx xxx Research report The effects of prime visibility on ERP measures of masked priming Phillip J. Holcomb a, T, Lindsay Reder
BRESC-40606; No. of pages: 18; 4C: DTD 5 Cognitive Brain Research xx (2005) xxx xxx Research report The effects of prime visibility on ERP measures of masked priming Phillip J. Holcomb a, T, Lindsay Reder
Neuroscience Letters
 Neuroscience Letters 469 (2010) 370 374 Contents lists available at ScienceDirect Neuroscience Letters journal homepage: www.elsevier.com/locate/neulet The influence on cognitive processing from the switches
Neuroscience Letters 469 (2010) 370 374 Contents lists available at ScienceDirect Neuroscience Letters journal homepage: www.elsevier.com/locate/neulet The influence on cognitive processing from the switches
Untangling syntactic and sensory processing: An ERP study of music perception
 Psychophysiology, 44 (2007), 476 490. Blackwell Publishing Inc. Printed in the USA. Copyright r 2007 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2007.00517.x Untangling syntactic
Psychophysiology, 44 (2007), 476 490. Blackwell Publishing Inc. Printed in the USA. Copyright r 2007 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2007.00517.x Untangling syntactic
This is an electronic reprint of the original article. This reprint may differ from the original in pagination and typographic detail.
 This is an electronic reprint of the original article. This reprint may differ from the original in pagination and typographic detail. Author(s): Maidhof, Clemens; Pitkäniemi, Anni; Tervaniemi, Mari Title:
This is an electronic reprint of the original article. This reprint may differ from the original in pagination and typographic detail. Author(s): Maidhof, Clemens; Pitkäniemi, Anni; Tervaniemi, Mari Title:
Proceedings of Meetings on Acoustics
 Proceedings of Meetings on Acoustics Volume 19, 2013 http://acousticalsociety.org/ ICA 2013 Montreal Montreal, Canada 2-7 June 2013 Psychological and Physiological Acoustics Session 4aPPb: Binaural Hearing
Proceedings of Meetings on Acoustics Volume 19, 2013 http://acousticalsociety.org/ ICA 2013 Montreal Montreal, Canada 2-7 June 2013 Psychological and Physiological Acoustics Session 4aPPb: Binaural Hearing
EEG Eye-Blinking Artefacts Power Spectrum Analysis
 EEG Eye-Blinking Artefacts Power Spectrum Analysis Plamen Manoilov Abstract: Artefacts are noises introduced to the electroencephalogram s (EEG) signal by not central nervous system (CNS) sources of electric
EEG Eye-Blinking Artefacts Power Spectrum Analysis Plamen Manoilov Abstract: Artefacts are noises introduced to the electroencephalogram s (EEG) signal by not central nervous system (CNS) sources of electric
ARTICLE IN PRESS. Neuroscience Letters xxx (2014) xxx xxx. Contents lists available at ScienceDirect. Neuroscience Letters
 xxx xxx. Contents lists available at ScienceDirect. Neuroscience Letters") NSL 30787 5 Neuroscience Letters xxx (204) xxx xxx Contents lists available at ScienceDirect Neuroscience Letters jo ur nal ho me page: www.elsevier.com/locate/neulet 2 3 4 Q 5 6 Earlier timbre processing
NSL 30787 5 Neuroscience Letters xxx (204) xxx xxx Contents lists available at ScienceDirect Neuroscience Letters jo ur nal ho me page: www.elsevier.com/locate/neulet 2 3 4 Q 5 6 Earlier timbre processing
CTP 431 Music and Audio Computing. Basic Acoustics. Graduate School of Culture Technology (GSCT) Juhan Nam
 Juhan Nam") CTP 431 Music and Audio Computing Basic Acoustics Graduate School of Culture Technology (GSCT) Juhan Nam 1 Outlines What is sound? Generation Propagation Reception Sound properties Loudness Pitch Timbre
CTP 431 Music and Audio Computing Basic Acoustics Graduate School of Culture Technology (GSCT) Juhan Nam 1 Outlines What is sound? Generation Propagation Reception Sound properties Loudness Pitch Timbre
PRODUCT SHEET
 ERS100C EVOKED RESPONSE AMPLIFIER MODULE The evoked response amplifier module (ERS100C) is a single channel, high gain, extremely low noise, differential input, biopotential amplifier designed to accurately
ERS100C EVOKED RESPONSE AMPLIFIER MODULE The evoked response amplifier module (ERS100C) is a single channel, high gain, extremely low noise, differential input, biopotential amplifier designed to accurately
Grand Rounds 5/15/2012
 Grand Rounds 5/15/2012 Department of Neurology P Dr. John Shelley-Tremblay, USA Psychology P I have no financial disclosures P I discuss no medications nore off-label uses of medications An Introduction
Grand Rounds 5/15/2012 Department of Neurology P Dr. John Shelley-Tremblay, USA Psychology P I have no financial disclosures P I discuss no medications nore off-label uses of medications An Introduction
Detection and correction of artefacts in EEG for neurofeedback and BCI applications
 Eindhoven University of Technology MASTER Detection and correction of artefacts in EEG for neurofeedback and BCI applications Erkens, I.J.M. Award date: 22 Disclaimer This document contains a student thesis
Eindhoven University of Technology MASTER Detection and correction of artefacts in EEG for neurofeedback and BCI applications Erkens, I.J.M. Award date: 22 Disclaimer This document contains a student thesis
Musical Acoustics Lecture 15 Pitch & Frequency (Psycho-Acoustics)
") 1 Musical Acoustics Lecture 15 Pitch & Frequency (Psycho-Acoustics) Pitch Pitch is a subjective characteristic of sound Some listeners even assign pitch differently depending upon whether the sound was
1 Musical Acoustics Lecture 15 Pitch & Frequency (Psycho-Acoustics) Pitch Pitch is a subjective characteristic of sound Some listeners even assign pitch differently depending upon whether the sound was
Noise evaluation based on loudness-perception characteristics of older adults
 Noise evaluation based on loudness-perception characteristics of older adults Kenji KURAKATA 1 ; Tazu MIZUNAMI 2 National Institute of Advanced Industrial Science and Technology (AIST), Japan ABSTRACT
Noise evaluation based on loudness-perception characteristics of older adults Kenji KURAKATA 1 ; Tazu MIZUNAMI 2 National Institute of Advanced Industrial Science and Technology (AIST), Japan ABSTRACT
Do Zwicker Tones Evoke a Musical Pitch?
 Do Zwicker Tones Evoke a Musical Pitch? Hedwig E. Gockel and Robert P. Carlyon Abstract It has been argued that musical pitch, i.e. pitch in its strictest sense, requires phase locking at the level of
Do Zwicker Tones Evoke a Musical Pitch? Hedwig E. Gockel and Robert P. Carlyon Abstract It has been argued that musical pitch, i.e. pitch in its strictest sense, requires phase locking at the level of
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland
 AWARD NUMBER: W81XWH-13-1-0491 TITLE: Default, Cognitive, and Affective Brain Networks in Human Tinnitus PRINCIPAL INVESTIGATOR: Jennifer R. Melcher, PhD CONTRACTING ORGANIZATION: Massachusetts Eye and
AWARD NUMBER: W81XWH-13-1-0491 TITLE: Default, Cognitive, and Affective Brain Networks in Human Tinnitus PRINCIPAL INVESTIGATOR: Jennifer R. Melcher, PhD CONTRACTING ORGANIZATION: Massachusetts Eye and
Abnormal Electrical Brain Responses to Pitch in Congenital Amusia Isabelle Peretz, PhD, 1 Elvira Brattico, MA, 2 and Mari Tervaniemi, PhD 2
 Abnormal Electrical Brain Responses to Pitch in Congenital Amusia Isabelle Peretz, PhD, 1 Elvira Brattico, MA, 2 and Mari Tervaniemi, PhD 2 Congenital amusia is a lifelong disability that prevents afflicted
Abnormal Electrical Brain Responses to Pitch in Congenital Amusia Isabelle Peretz, PhD, 1 Elvira Brattico, MA, 2 and Mari Tervaniemi, PhD 2 Congenital amusia is a lifelong disability that prevents afflicted
THE INTERACTION BETWEEN MELODIC PITCH CONTENT AND RHYTHMIC PERCEPTION. Gideon Broshy, Leah Latterner and Kevin Sherwin
 THE INTERACTION BETWEEN MELODIC PITCH CONTENT AND RHYTHMIC PERCEPTION. BACKGROUND AND AIMS [Leah Latterner]. Introduction Gideon Broshy, Leah Latterner and Kevin Sherwin Yale University, Cognition of Musical
THE INTERACTION BETWEEN MELODIC PITCH CONTENT AND RHYTHMIC PERCEPTION. BACKGROUND AND AIMS [Leah Latterner]. Introduction Gideon Broshy, Leah Latterner and Kevin Sherwin Yale University, Cognition of Musical
NeXus: Event-Related potentials Evoked potentials for Psychophysiology & Neuroscience
 NeXus: Event-Related potentials Evoked potentials for Psychophysiology & Neuroscience This NeXus white paper has been created to educate and inform the reader about the Event Related Potentials (ERP) and
NeXus: Event-Related potentials Evoked potentials for Psychophysiology & Neuroscience This NeXus white paper has been created to educate and inform the reader about the Event Related Potentials (ERP) and
Informational masking of speech produced by speech-like sounds without linguistic content
 Informational masking of speech produced by speech-like sounds without linguistic content Jing Chen, Huahui Li, Liang Li, and Xihong Wu a) Department of Machine Intelligence, Speech and Hearing Research
Informational masking of speech produced by speech-like sounds without linguistic content Jing Chen, Huahui Li, Liang Li, and Xihong Wu a) Department of Machine Intelligence, Speech and Hearing Research
Communicating hands: ERPs elicited by meaningful symbolic hand postures
 Neuroscience Letters 372 (2004) 52 56 Communicating hands: ERPs elicited by meaningful symbolic hand postures Thomas C. Gunter a,, Patric Bach b a Max-Planck-Institute for Human Cognitive and Brain Sciences,
Neuroscience Letters 372 (2004) 52 56 Communicating hands: ERPs elicited by meaningful symbolic hand postures Thomas C. Gunter a,, Patric Bach b a Max-Planck-Institute for Human Cognitive and Brain Sciences,
MASTER'S THESIS. Listener Envelopment
 MASTER'S THESIS 2008:095 Listener Envelopment Effects of changing the sidewall material in a model of an existing concert hall Dan Nyberg Luleå University of Technology Master thesis Audio Technology Department
MASTER'S THESIS 2008:095 Listener Envelopment Effects of changing the sidewall material in a model of an existing concert hall Dan Nyberg Luleå University of Technology Master thesis Audio Technology Department
Experiments on tone adjustments
 Experiments on tone adjustments Jesko L. VERHEY 1 ; Jan HOTS 2 1 University of Magdeburg, Germany ABSTRACT Many technical sounds contain tonal components originating from rotating parts, such as electric
Experiments on tone adjustments Jesko L. VERHEY 1 ; Jan HOTS 2 1 University of Magdeburg, Germany ABSTRACT Many technical sounds contain tonal components originating from rotating parts, such as electric
The Research of Controlling Loudness in the Timbre Subjective Perception Experiment of Sheng
 The Research of Controlling Loudness in the Timbre Subjective Perception Experiment of Sheng S. Zhu, P. Ji, W. Kuang and J. Yang Institute of Acoustics, CAS, O.21, Bei-Si-huan-Xi Road, 100190 Beijing,
The Research of Controlling Loudness in the Timbre Subjective Perception Experiment of Sheng S. Zhu, P. Ji, W. Kuang and J. Yang Institute of Acoustics, CAS, O.21, Bei-Si-huan-Xi Road, 100190 Beijing,
Hearing Research 327 (2015) 9e27. Contents lists available at ScienceDirect. Hearing Research. journal homepage:
 9e27. Contents lists available at ScienceDirect. Hearing Research. journal homepage:") Hearing Research 327 (2015) 9e27 Contents lists available at ScienceDirect Hearing Research journal homepage: www.elsevier.com/locate/heares Research paper Evidence for differential modulation of primary
Hearing Research 327 (2015) 9e27 Contents lists available at ScienceDirect Hearing Research journal homepage: www.elsevier.com/locate/heares Research paper Evidence for differential modulation of primary
Supplemental Material for Gamma-band Synchronization in the Macaque Hippocampus and Memory Formation
 Supplemental Material for Gamma-band Synchronization in the Macaque Hippocampus and Memory Formation Michael J. Jutras, Pascal Fries, Elizabeth A. Buffalo * *To whom correspondence should be addressed.
Supplemental Material for Gamma-band Synchronization in the Macaque Hippocampus and Memory Formation Michael J. Jutras, Pascal Fries, Elizabeth A. Buffalo * *To whom correspondence should be addressed.
The perception of concurrent sound objects through the use of harmonic enhancement: a study of auditory attention
 Atten Percept Psychophys (2015) 77:922 929 DOI 10.3758/s13414-014-0826-9 The perception of concurrent sound objects through the use of harmonic enhancement: a study of auditory attention Elena Koulaguina
Atten Percept Psychophys (2015) 77:922 929 DOI 10.3758/s13414-014-0826-9 The perception of concurrent sound objects through the use of harmonic enhancement: a study of auditory attention Elena Koulaguina
Brian C. J. Moore Department of Experimental Psychology, University of Cambridge, Downing Street, Cambridge CB2 3EB, England
 Asymmetry of masking between complex tones and noise: Partial loudness Hedwig Gockel a) CNBH, Department of Physiology, University of Cambridge, Downing Street, Cambridge CB2 3EG, England Brian C. J. Moore
Asymmetry of masking between complex tones and noise: Partial loudness Hedwig Gockel a) CNBH, Department of Physiology, University of Cambridge, Downing Street, Cambridge CB2 3EG, England Brian C. J. Moore
Effect of room acoustic conditions on masking efficiency
 Effect of room acoustic conditions on masking efficiency Hyojin Lee a, Graduate school, The University of Tokyo Komaba 4-6-1, Meguro-ku, Tokyo, 153-855, JAPAN Kanako Ueno b, Meiji University, JAPAN Higasimita
Effect of room acoustic conditions on masking efficiency Hyojin Lee a, Graduate school, The University of Tokyo Komaba 4-6-1, Meguro-ku, Tokyo, 153-855, JAPAN Kanako Ueno b, Meiji University, JAPAN Higasimita
Acoustic Prosodic Features In Sarcastic Utterances
 Acoustic Prosodic Features In Sarcastic Utterances Introduction: The main goal of this study is to determine if sarcasm can be detected through the analysis of prosodic cues or acoustic features automatically.
Acoustic Prosodic Features In Sarcastic Utterances Introduction: The main goal of this study is to determine if sarcasm can be detected through the analysis of prosodic cues or acoustic features automatically.
The Time Course of Orthographic and Phonological Code Activation Jonathan Grainger, 1 Kristi Kiyonaga, 2 and Phillip J. Holcomb 2
 PSYCHOLOGICAL SCIENCE Research Report The Time Course of Orthographic and Phonological Code Activation Jonathan Grainger, 1 Kristi Kiyonaga, 2 and Phillip J. Holcomb 2 1 CNRS and University of Provence,
PSYCHOLOGICAL SCIENCE Research Report The Time Course of Orthographic and Phonological Code Activation Jonathan Grainger, 1 Kristi Kiyonaga, 2 and Phillip J. Holcomb 2 1 CNRS and University of Provence,
Melodic pitch expectation interacts with neural responses to syntactic but not semantic violations
 cortex xxx () e Available online at www.sciencedirect.com Journal homepage: www.elsevier.com/locate/cortex Research report Melodic pitch expectation interacts with neural responses to syntactic but not
cortex xxx () e Available online at www.sciencedirect.com Journal homepage: www.elsevier.com/locate/cortex Research report Melodic pitch expectation interacts with neural responses to syntactic but not
MUSICAL EAR TRAINING THROUGH ACTIVE MUSIC MAKING IN ADOLESCENT Cl USERS. The background ~
 It's good news that more and more teenagers are being offered the option of cochlear implants. They are candidates who require information and support given in a way to meet their particular needs which
It's good news that more and more teenagers are being offered the option of cochlear implants. They are candidates who require information and support given in a way to meet their particular needs which
LOUDNESS EFFECT OF THE DIFFERENT TONES ON THE TIMBRE SUBJECTIVE PERCEPTION EXPERIMENT OF ERHU
 The 21 st International Congress on Sound and Vibration 13-17 July, 2014, Beijing/China LOUDNESS EFFECT OF THE DIFFERENT TONES ON THE TIMBRE SUBJECTIVE PERCEPTION EXPERIMENT OF ERHU Siyu Zhu, Peifeng Ji,
The 21 st International Congress on Sound and Vibration 13-17 July, 2014, Beijing/China LOUDNESS EFFECT OF THE DIFFERENT TONES ON THE TIMBRE SUBJECTIVE PERCEPTION EXPERIMENT OF ERHU Siyu Zhu, Peifeng Ji,
Ellen F. Lau 1,2,3. Phillip J. Holcomb 2. Gina R. Kuperberg 1,2
 DISSOCIATING N400 EFFECTS OF PREDICTION FROM ASSOCIATION IN SINGLE WORD CONTEXTS Ellen F. Lau 1,2,3 Phillip J. Holcomb 2 Gina R. Kuperberg 1,2 1 Athinoula C. Martinos Center for Biomedical Imaging, Massachusetts
DISSOCIATING N400 EFFECTS OF PREDICTION FROM ASSOCIATION IN SINGLE WORD CONTEXTS Ellen F. Lau 1,2,3 Phillip J. Holcomb 2 Gina R. Kuperberg 1,2 1 Athinoula C. Martinos Center for Biomedical Imaging, Massachusetts
Cross-modal Semantic Priming: A Timecourse Analysis Using Event-related Brain Potentials
 LANGUAGE AND COGNITIVE PROCESSES, 1993, 8 (4) 379-411 Cross-modal Semantic Priming: A Timecourse Analysis Using Event-related Brain Potentials Phillip J. Holcomb and Jane E. Anderson Department of Psychology,
LANGUAGE AND COGNITIVE PROCESSES, 1993, 8 (4) 379-411 Cross-modal Semantic Priming: A Timecourse Analysis Using Event-related Brain Potentials Phillip J. Holcomb and Jane E. Anderson Department of Psychology,
Frequency and predictability effects on event-related potentials during reading
 Research Report Frequency and predictability effects on event-related potentials during reading Michael Dambacher a,, Reinhold Kliegl a, Markus Hofmann b, Arthur M. Jacobs b a Helmholtz Center for the
Research Report Frequency and predictability effects on event-related potentials during reading Michael Dambacher a,, Reinhold Kliegl a, Markus Hofmann b, Arthur M. Jacobs b a Helmholtz Center for the
Information processing in high- and low-risk parents: What can we learn from EEG?
 Information processing in high- and low-risk parents: What can we learn from EEG? Social Information Processing What differentiates parents who abuse their children from parents who don t? Mandy M. Rabenhorst
Information processing in high- and low-risk parents: What can we learn from EEG? Social Information Processing What differentiates parents who abuse their children from parents who don t? Mandy M. Rabenhorst
Individual differences in prediction: An investigation of the N400 in word-pair semantic priming
 Individual differences in prediction: An investigation of the N400 in word-pair semantic priming Xiao Yang & Lauren Covey Cognitive and Brain Sciences Brown Bag Talk October 17, 2016 Caitlin Coughlin,
Individual differences in prediction: An investigation of the N400 in word-pair semantic priming Xiao Yang & Lauren Covey Cognitive and Brain Sciences Brown Bag Talk October 17, 2016 Caitlin Coughlin,
The Processing of Pitch and Scale: An ERP Study of Musicians Trained Outside of the Western Musical System
 The Processing of Pitch and Scale: An ERP Study of Musicians Trained Outside of the Western Musical System LAURA BISCHOFF RENNINGER [1] Shepherd University MICHAEL P. WILSON University of Illinois EMANUEL
The Processing of Pitch and Scale: An ERP Study of Musicians Trained Outside of the Western Musical System LAURA BISCHOFF RENNINGER [1] Shepherd University MICHAEL P. WILSON University of Illinois EMANUEL
Event-Related Brain Potentials (ERPs) Elicited by Novel Stimuli during Sentence Processing
 Elicited by Novel Stimuli during Sentence Processing") Event-Related Brain Potentials (ERPs) Elicited by Novel Stimuli during Sentence Processing MARTA KUTAS AND STEVEN A. HILLYARD Department of Neurosciences School of Medicine University of California at
Event-Related Brain Potentials (ERPs) Elicited by Novel Stimuli during Sentence Processing MARTA KUTAS AND STEVEN A. HILLYARD Department of Neurosciences School of Medicine University of California at
Reconstruction of Ca 2+ dynamics from low frame rate Ca 2+ imaging data CS229 final project. Submitted by: Limor Bursztyn
 Reconstruction of Ca 2+ dynamics from low frame rate Ca 2+ imaging data CS229 final project. Submitted by: Limor Bursztyn Introduction Active neurons communicate by action potential firing (spikes), accompanied
Reconstruction of Ca 2+ dynamics from low frame rate Ca 2+ imaging data CS229 final project. Submitted by: Limor Bursztyn Introduction Active neurons communicate by action potential firing (spikes), accompanied
I like my coffee with cream and sugar. I like my coffee with cream and socks. I shaved off my mustache and beard. I shaved off my mustache and BEARD
 I like my coffee with cream and sugar. I like my coffee with cream and socks I shaved off my mustache and beard. I shaved off my mustache and BEARD All turtles have four legs All turtles have four leg
I like my coffee with cream and sugar. I like my coffee with cream and socks I shaved off my mustache and beard. I shaved off my mustache and BEARD All turtles have four legs All turtles have four leg
Topic 10. Multi-pitch Analysis
 Topic 10 Multi-pitch Analysis What is pitch? Common elements of music are pitch, rhythm, dynamics, and the sonic qualities of timbre and texture. An auditory perceptual attribute in terms of which sounds
Topic 10 Multi-pitch Analysis What is pitch? Common elements of music are pitch, rhythm, dynamics, and the sonic qualities of timbre and texture. An auditory perceptual attribute in terms of which sounds
Neural evidence for a single lexicogrammatical processing system. Jennifer Hughes
 Neural evidence for a single lexicogrammatical processing system Jennifer Hughes j.j.hughes@lancaster.ac.uk Background Approaches to collocation Background Association measures Background EEG, ERPs, and
Neural evidence for a single lexicogrammatical processing system Jennifer Hughes j.j.hughes@lancaster.ac.uk Background Approaches to collocation Background Association measures Background EEG, ERPs, and
2018 Fall CTP431: Music and Audio Computing Fundamentals of Musical Acoustics
 2018 Fall CTP431: Music and Audio Computing Fundamentals of Musical Acoustics Graduate School of Culture Technology, KAIST Juhan Nam Outlines Introduction to musical tones Musical tone generation - String
2018 Fall CTP431: Music and Audio Computing Fundamentals of Musical Acoustics Graduate School of Culture Technology, KAIST Juhan Nam Outlines Introduction to musical tones Musical tone generation - String
German Center for Music Therapy Research
 Effects of music therapy for adult CI users on the perception of music, prosody in speech, subjective self-concept and psychophysiological arousal Research Network: E. Hutter, M. Grapp, H. Argstatter,
Effects of music therapy for adult CI users on the perception of music, prosody in speech, subjective self-concept and psychophysiological arousal Research Network: E. Hutter, M. Grapp, H. Argstatter,
Affective Priming. Music 451A Final Project
 Affective Priming Music 451A Final Project The Question Music often makes us feel a certain way. Does this feeling have semantic meaning like the words happy or sad do? Does music convey semantic emotional
Affective Priming Music 451A Final Project The Question Music often makes us feel a certain way. Does this feeling have semantic meaning like the words happy or sad do? Does music convey semantic emotional
EFFECT OF REPETITION OF STANDARD AND COMPARISON TONES ON RECOGNITION MEMORY FOR PITCH '
 Journal oj Experimental Psychology 1972, Vol. 93, No. 1, 156-162 EFFECT OF REPETITION OF STANDARD AND COMPARISON TONES ON RECOGNITION MEMORY FOR PITCH ' DIANA DEUTSCH " Center for Human Information Processing,
Journal oj Experimental Psychology 1972, Vol. 93, No. 1, 156-162 EFFECT OF REPETITION OF STANDARD AND COMPARISON TONES ON RECOGNITION MEMORY FOR PITCH ' DIANA DEUTSCH " Center for Human Information Processing,
The Effects of Study Condition Preference on Memory and Free Recall LIANA, MARISSA, JESSI AND BROOKE
 The Effects of Study Condition Preference on Memory and Free Recall LIANA, MARISSA, JESSI AND BROOKE Introduction -Salamè & Baddeley 1988 Presented nine digits on a computer screen for 750 milliseconds
The Effects of Study Condition Preference on Memory and Free Recall LIANA, MARISSA, JESSI AND BROOKE Introduction -Salamè & Baddeley 1988 Presented nine digits on a computer screen for 750 milliseconds
Blending in action: Diagrams reveal conceptual integration in routine activity
 Cognitive Science Online, Vol.1, pp.34 45, 2003 http://cogsci-online.ucsd.edu Blending in action: Diagrams reveal conceptual integration in routine activity Beate Schwichtenberg Department of Cognitive
Cognitive Science Online, Vol.1, pp.34 45, 2003 http://cogsci-online.ucsd.edu Blending in action: Diagrams reveal conceptual integration in routine activity Beate Schwichtenberg Department of Cognitive
SHORT TERM PITCH MEMORY IN WESTERN vs. OTHER EQUAL TEMPERAMENT TUNING SYSTEMS
 SHORT TERM PITCH MEMORY IN WESTERN vs. OTHER EQUAL TEMPERAMENT TUNING SYSTEMS Areti Andreopoulou Music and Audio Research Laboratory New York University, New York, USA aa1510@nyu.edu Morwaread Farbood
SHORT TERM PITCH MEMORY IN WESTERN vs. OTHER EQUAL TEMPERAMENT TUNING SYSTEMS Areti Andreopoulou Music and Audio Research Laboratory New York University, New York, USA aa1510@nyu.edu Morwaread Farbood
Event-Related Brain Potentials Reflect Semantic Priming in an Object Decision Task
 BRAIN AND COGNITION 24, 259-276 (1994) Event-Related Brain Potentials Reflect Semantic Priming in an Object Decision Task PHILLIP.1. HOLCOMB AND WARREN B. MCPHERSON Tufts University Subjects made speeded
BRAIN AND COGNITION 24, 259-276 (1994) Event-Related Brain Potentials Reflect Semantic Priming in an Object Decision Task PHILLIP.1. HOLCOMB AND WARREN B. MCPHERSON Tufts University Subjects made speeded
VivoSense. User Manual Galvanic Skin Response (GSR) Analysis Module. VivoSense, Inc. Newport Beach, CA, USA Tel. (858) , Fax.
 Analysis Module. VivoSense, Inc. Newport Beach, CA, USA Tel. (858) , Fax.") VivoSense User Manual Galvanic Skin Response (GSR) Analysis VivoSense Version 3.1 VivoSense, Inc. Newport Beach, CA, USA Tel. (858) 876-8486, Fax. (248) 692-0980 Email: info@vivosense.com; Web: www.vivosense.com
VivoSense User Manual Galvanic Skin Response (GSR) Analysis VivoSense Version 3.1 VivoSense, Inc. Newport Beach, CA, USA Tel. (858) 876-8486, Fax. (248) 692-0980 Email: info@vivosense.com; Web: www.vivosense.com
Auditory semantic networks for words and natural sounds
 available at www.sciencedirect.com www.elsevier.com/locate/brainres Research Report Auditory semantic networks for words and natural sounds A. Cummings a,b,c,,r.čeponienė a, A. Koyama a, A.P. Saygin c,f,
available at www.sciencedirect.com www.elsevier.com/locate/brainres Research Report Auditory semantic networks for words and natural sounds A. Cummings a,b,c,,r.čeponienė a, A. Koyama a, A.P. Saygin c,f,
On the locus of the semantic satiation effect: Evidence from event-related brain potentials
 Memory & Cognition 2000, 28 (8), 1366-1377 On the locus of the semantic satiation effect: Evidence from event-related brain potentials JOHN KOUNIOS University of Pennsylvania, Philadelphia, Pennsylvania
Memory & Cognition 2000, 28 (8), 1366-1377 On the locus of the semantic satiation effect: Evidence from event-related brain potentials JOHN KOUNIOS University of Pennsylvania, Philadelphia, Pennsylvania
Why are natural sounds detected faster than pips?
 Why are natural sounds detected faster than pips? Clara Suied Department of Physiology, Development and Neuroscience, Centre for the Neural Basis of Hearing, Downing Street, Cambridge CB2 3EG, United Kingdom
Why are natural sounds detected faster than pips? Clara Suied Department of Physiology, Development and Neuroscience, Centre for the Neural Basis of Hearing, Downing Street, Cambridge CB2 3EG, United Kingdom
Music Training and Neuroplasticity
 Presents Music Training and Neuroplasticity Searching For the Mind with John Leif, M.D. Neuroplasticity... 2 The brain's ability to reorganize itself by forming new neural connections throughout life....
Presents Music Training and Neuroplasticity Searching For the Mind with John Leif, M.D. Neuroplasticity... 2 The brain's ability to reorganize itself by forming new neural connections throughout life....
Processing Linguistic and Musical Pitch by English-Speaking Musicians and Non-Musicians
 Proceedings of the 20th North American Conference on Chinese Linguistics (NACCL-20). 2008. Volume 1. Edited by Marjorie K.M. Chan and Hana Kang. Columbus, Ohio: The Ohio State University. Pages 139-145.
Proceedings of the 20th North American Conference on Chinese Linguistics (NACCL-20). 2008. Volume 1. Edited by Marjorie K.M. Chan and Hana Kang. Columbus, Ohio: The Ohio State University. Pages 139-145.
Nature Neuroscience: doi: /nn Supplementary Figure 1. Emergence of dmpfc and BLA 4-Hz oscillations during freezing behavior.
 Supplementary Figure 1 Emergence of dmpfc and BLA 4-Hz oscillations during freezing behavior. (a) Representative power spectrum of dmpfc LFPs recorded during Retrieval for freezing and no freezing periods.
Supplementary Figure 1 Emergence of dmpfc and BLA 4-Hz oscillations during freezing behavior. (a) Representative power spectrum of dmpfc LFPs recorded during Retrieval for freezing and no freezing periods.
An ERP study of low and high relevance semantic features
 Brain Research Bulletin 69 (2006) 182 186 An ERP study of low and high relevance semantic features Giuseppe Sartori a,, Francesca Mameli a, David Polezzi a, Luigi Lombardi b a Department of General Psychology,
Brain Research Bulletin 69 (2006) 182 186 An ERP study of low and high relevance semantic features Giuseppe Sartori a,, Francesca Mameli a, David Polezzi a, Luigi Lombardi b a Department of General Psychology,
Estimating the Time to Reach a Target Frequency in Singing
 THE NEUROSCIENCES AND MUSIC III: DISORDERS AND PLASTICITY Estimating the Time to Reach a Target Frequency in Singing Sean Hutchins a and David Campbell b a Department of Psychology, McGill University,
THE NEUROSCIENCES AND MUSIC III: DISORDERS AND PLASTICITY Estimating the Time to Reach a Target Frequency in Singing Sean Hutchins a and David Campbell b a Department of Psychology, McGill University,
Modeling sound quality from psychoacoustic measures
 Modeling sound quality from psychoacoustic measures Lena SCHELL-MAJOOR 1 ; Jan RENNIES 2 ; Stephan D. EWERT 3 ; Birger KOLLMEIER 4 1,2,4 Fraunhofer IDMT, Hör-, Sprach- und Audiotechnologie & Cluster of
Modeling sound quality from psychoacoustic measures Lena SCHELL-MAJOOR 1 ; Jan RENNIES 2 ; Stephan D. EWERT 3 ; Birger KOLLMEIER 4 1,2,4 Fraunhofer IDMT, Hör-, Sprach- und Audiotechnologie & Cluster of
With thanks to Seana Coulson and Katherine De Long!
 Event Related Potentials (ERPs): A window onto the timing of cognition Kim Sweeney COGS1- Introduction to Cognitive Science November 19, 2009 With thanks to Seana Coulson and Katherine De Long! Overview
Event Related Potentials (ERPs): A window onto the timing of cognition Kim Sweeney COGS1- Introduction to Cognitive Science November 19, 2009 With thanks to Seana Coulson and Katherine De Long! Overview